
نگاهی دوباره بر پیچیدگیکاهشناپذیرویلیام دمبسکی/ ترجمه: امیرحسین لطیفی

درآمد
مفهوم پیچیدگیکاهشناپذیر[1] مایکل بیهی، و به طور خاص استفادهی او از این مفهوم برای نقد داروینیسم، هنوز هم که هنوز است زیر رگبار آتش محافل زیستشناسان است. مشکل بیهی، آنطور که داروینیستها میگویند، این است که مشکلی درست کرده که در واقع مشکل نیست و بدون این که مانعی در مقابل مکانیسم تغییر تصادفی داروین و انتخابطبیعی ایجاد کند، با همین مکانیسم قابل توضیح است. اما آیا واقعا چنین است؟ هشت سال از زمانی که بیهی پیچیدگیکاهشناپذیر را در کتاب جعبه سیاه داروین[2] خود معرفی کرد، میگذرد و این کتاب همچنان هر ساله 15000 نسخه فقط به زبان انگلیسی میفروشد. در این مقاله قصد دارم نگاهی دوباره به مفهوم پیچیدگیکاهشناپذیر بیهی بیندازم و نشان دهم چرا معضلی که او ایجاد کرده، حالا حتی از زمانی که او برای او نخستین بار این مفهوم را مطرح کرد، برای داروینیستها آزاردهندهتر است. چهار بخش اول این مقاله مرور و بسطی است بر مطالبی که من پیشتر در جاهای دیگر مطرح کردهام. بخش آخر اما مطالب جدیدی دارد.
۱- تعریف پیچیدگیکاهشناپذیر
ماشینهای مولکولی بسیار پیچیده نقشی بنیادی در حیات سلول بازی میکنند و هر روز توجهات بیشتری را در مجامع زیستشناسی به خود جلب میکنند. برا/ی نمونه، در فوریهی سال 1998 مجله زیستشناسی سل[3] یک شماره را منحصرا به «ماشینهای ماکرومولکولی» اختصاص داد. همهی سلولها از ماشینهای مولکولی پیچیده برای پردازش اطلاعات، تبدیل انرژی، سوخت و ساز مواد معدنی، ساخت پروتئینها و تبادل مواد بین غشا استفاده میکنند. بروس آلبرتز[4]، رییس آکادمی ملی علوم[5]، این شماره را با مقالهای باعنوان سلول به مثابه مجموعهای از ماشینهای پروتئینی[6] معرفی کرد. او در این مقاله مینویسد:
ما همواره سلول را دست کم میگیریم . . . کل سلول را میتوان به عنوان کارخانهای در نظر گرفت که شامل شبکهای بینظیر از خطوط مونتاژ به هم پیوسته است که هر کدام از مجموعهای از ماشینهای پروتئینی بزرگ ساخته شدهاند . . . چرا این پروتئینهای بزرگ مونتاژکننده را که بنیان عملکرد سلول را تشکیل میدهند، ماشینهای پروتئینی مینامیم؟ چون دقیقا مانند ماشینهایی که انسان اختراع کرده تا به طرزی کارآمد به امور مربوط به جهان ماکروسکوپی بپردازند، این پروتئینها هم شامل بخشهای حرکتیاند و بسیار هماهنگ کار میکنند]1[.
تقریبا شش سال بعد، دسامبر 2003، مجلهی بایو اسیز[7] شمارهای اختصاصی دربارهی «ماشینهای مولکولی» را منتشر کرد. در مقالهی معرفی این شماره، آدام ویلکینز[8]، ویراستار بایو اسیز مینویسد:
مقالاتی که این شماره در بردارد برخی شباهتهای قابل توجه را میان ماشینهای مولکولی/زیستی و ماشینهای دستساز بشر را به تصویر میکشد. نخست این که ماشینهای مولکولی مانند ماشینهای دستساز بشر، عملکردهای بسیار تخصصیای انجام میدهند. ثانیا، مجموعههای ماکرومولکولی بخشهای متعددی را نشان میدهند که این بخشها به شیوهای دقیق و معین با همدیگر تعامل میکنند و ورودی و خروجیهای کاملا مشخصی دارند. سوم این که بسیاری از این ماشینها بخشهایی دارند که میتوانند در ماشینهای مولکولی دیگر هم (نهایتا با اندکی تغییرات) مورد استفاده قرار گیرند که با بخشهای قابل تعویض ماشینهای مصنوعی، قابل مقایسه است. در نهایت، که مطمئنا آخری نیست، ماشینهای زیستی اساسیترین ویژگی ماشینها را دارند؛ یعنی همگی آنها انرژی را به «کار» تبدیل میکنند]2[.
آلبرتز و ویلکینز توجهات را به شباهت فراوان میان ماشینهای مولکولی و ماشینهایی که به دست مهندسی انسانی طراحیشدهاند، جلب میکنند. با این حال، نوداروینیها پیچیدگیهای حیرتانگیز سلول را محصولات فرگشت داروینی در نظرمیگیرند و معتقدند که آنها «به نظر میرسد» طراحی شدهاند. در دههی 1990 اما دانشمندان چالشی را با نظریهی نوداروینی آغاز کردند. آنها استدلالشان این بود که چنین ماشینهای پروتئینیای تنها میتوانند با یک طراحی واقعی شکل گرفته باشند. برای مثال، در سال 1996 زیستشیمیدان دانشگاه لی[9]، مایکل بیهی، کتابی با عنوان جعبه سیاه داروین[10] منتشر کرد. در این کتاب او به تفصیل ناکامیهای نظریهی نوداروینی در مورد منشا ماشینهای مولکولی پیچیده در سلول را بیان میکند. بیهی اما به این اکتفا نمیکند. او همچنین استدلال میکند که این ماشینهای مولکولی، طراحی واقعی را نشان میدهند. در مرکز استدلال او ایدهی «پیچیدگیکاهشناپذیر»[11] قرار داشت.
یک سیستم دارای عملکرد، زمانی یک مجموعهی کاهشناپذیر است که شامل چندین زیرمجموعه (برای مثال دو یا چند مجموعه از بخشهای نامرتبط) باشد که بدون از بین رفتن عملکرد اصلی سیستم نتوان آنها را ساده کرد. من برای این زیرمجموعههای چندبخشی از عنوان «هستهی کاهشناپذیر»[12]]3[ سیستم یاد میکنم. این تعریف از آنچه که در آغاز ممکن است به نظر برسد دقیقتر است، پس بیاید نگاهی از نزدیک به آن بیندازیم. سیستمهای پیچیدهی کاهشناپذیر به یک طبقهی وسیعتر به نام سیستمهای کارکردی یکپارچه تعلق دارند. این سیستمها شامل بخشهایی هستند که کاملا منطبق بر یکدیگرند و این موضوع عملکرد سیستم را نسبت به تغییرات مجزا در آن بخشها به شدت حساس میکند. برای یک سیستم یکپارچه، یک تغییر در یک مکان اغلب کل سیستم را از کار میاندازد یا چندین تغییر در جایی دیگر باید اتفاق بیفتد تا سیستم به عملکردش ادامه دهد. بدین ترتیب میتوانیم «هستهی» یک سیستم عملکردی یکپارچه را براساس آن بخشهایی که برای عملکرد اساسی سیستم ضروری هستند، این گونه تعریف کنیم: «اگر بخشهایی از هسته را حذف کنیم، دیگر نمیتوان عملکرد اصلی سیستم را از سایر بخشهای باقیمانده بازیابی کرد. وقتی میگوییم که هستهای «کاهشناپذیر» است، منظور این است که هیچ سیستم دیگری با هستههای سادهتر نمیتوانند عملکرد اصلی سیستم ما را اجرا کنند.
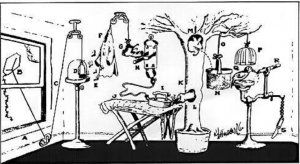
شکل۱- مدادتراش نخی گلدبرگ: «پنجره را باز کن (A) بادبادک را به هوا بفرست (B). نخ (C) در کوچک را بالا میبرد (D) اجازه میدهد تا بیدها (E) فرار کنند و پیرهن فلانل قرمز را بخورند(F) . در حالی که وزن پیراهن کم و کمتر میشود، کفش (G) دسته را میچرخاند (H) که باعث گرم شدن اتو میشود (I) و شلوار را میسوزاند (J). دود (K) وارد سوراخ روی درخت میشود (L) و سنجاب را بیرون میکشد (M) سنجاب درون سبد میپرد (N) و نخی را میکشد (O) که قفس را بالا میبرد(P) . بالا رفتن قفس به دارکوب (Q) اجازه میدهد تا مداد را بتراشد (R). چاقوی اضطراری (S) هم برای مواقعی که سنجاب یا دارکوب مریض بودند و نمیتوانستند کار کنند وجود دارد.» این یک سیستم عملکردی یک پارچه نیست!
عملکردهای اصلی یک سیستم از سه چیز تشکیل شده است: (1) کاری که سیستم به طور طبیعی و در بستر مناسبش انجام میدهد؛ این مورد به عنوان «عملکرد پایهای» یا «عملکرد اصلی» سیستم شناخته میشود. (2) حداقل عملکرد لازم تا سیستم کارکردی رضایتبخش در شرایط طبیعی یا بستر مناسبش داشته باشد؛ این را با عنوان «حداقل عملکرد» سیستم میشناسند. (3) راه و روشی که طی آن سیستم عملکرد پایهاش را اجرا میکند و به عنوان طریقهی عملکرد شناخته میشود. از آنجا که عملکرد پایهای شامل طرقی میشود که آن عملکرد صورت میگیرد، از این رو عملکرد پایهای صرفا با هدف سیستم سر و کار ندارد، بلکه به روشهای انجام شدن آن هم وابسته است. برای مثال، میخ و چسب، هر دو ممکن است که با یک عملکرد پایهای قطعات چوب را بهم متصل کنند و در یک بستر معین دقیقا یکسان عمل کنند، اما نحوهای که هر کدام عمل میکنند متفاوت است.
در همان راستا، موتور یک قایق موتوری را در نظر بگیرید که عملکرد پایهاش این است که یک قایق ماهیگیری کوچک را به دور یک دریاچه به حرکت درآورد و این کار را به کمک یک موتور الکتریکی یا بنزینی که پروانهای را میچرخاند، انجام میدهد. موتور قایق موتوری یک مجموعهی کاهشناپذیر است و هستهی تقلیل ناپذیرش، در کنار سایر بخشها شامل یک پروانه، یک موتور و میل گاردان است که موتور را به پروانه متصل میکند. حالا میتوانیم تصور کنیم این چیدمان را با جایگزین کردن یک بند کشی به جای موتور و میل گاردان، ساده کنیم. به طوری که کش با پیچیدن خودش پروانه را به چرخش در آورد. اما بعید به نظر میرسد چنین چینشی بازدهی کافی برای به حرکت درآوردن قایق به دور دریاچه را تامین کند. به عبارت دیگر، بسیار نامحتمل است که عملکرد حداقلی با بند کشی تامین شود. با این حال حتی اگر این عملکرد حداقلی هم تامین میشد، این چیدمان عملکرد پایهای را به همان صورتی که موتور قایق انجام میدهد، اجرا نمیکند: موتور اصلی به چرخش روتور (بخش چرخندهی موتور) متکی است نه پیچش یک محیط الاستیک.
به عنوان یک مثال دیگر از سیستمهای پیچیدهی کاهشناپذیر، یک ساعت جیبی قدیمی را فرض کنید. عملکرد پایهای ساعت این است که از طریق عملکرد چرخدندهها ساعت را اعلام کند. بخشهای مختلفی از ساعت برای عملکرد پایهی آن لازم و غیرقابل چشمپوشیاند؛ برای نمونه فنر، صفحه و عقربه. همهی اینها متعلق به هستهی کاهشناپذیرند. اما توجه داشته باشید که باقی قسمتهای ساعت قابل کنار گداشتناند. برای مثال، شیشه، قاب فلزی و زنجیر. این بخشها برای عملکرد پایهی ساعت غیرضروری و اضافیاند و به هستهی کاهشناپذیر تعلق ندارند. این که سایر قسمتها به هستهی کاهشناپذیر تعلق دارند یا نه، به حداقل سطح عملکرد مورد انتظار از ساعت بستگی دارد. عقربهی ساعتشمار به تنهایی برای گفتن ساعت یا حتی محدودهی معینی از دقایق کافی است. اما اگر برایمان مهم باشد که دقیقه را به طور دقیق بدانیم، آن وقت عقربهی دقیقهشمار هم در هستهی کاهشناپذیر ضروری میشود. توجه داشته باشید که بسیاری از سیستمهای پیچیدهی کاهشناپذیر مانند ساعت جیبی بخشهایی دارند که برای عملکرد پایهای سیستم حیاتی نیستند. اینها همان بخشهایی هستند که خارج از هستهی کاهشناپذیر سیستم جای میگیرند.
در یک سیستم پیچیدهی کاهشناپذیر هر کدام از بخشهای هستهی کاهشناپذیر نقشی غیرقابل چشمپوشی در دستیابی سیستم به عملکرد پایه دارند. بنابراین حذف هر بخش، حتی یک بخش منفرد از هستهی کاهشناپذیر منجر به از دست رفتن کامل عملکرد پایهای سیستم میشود. با وجود این، برای تعیین این که یک سیستم پیچیده کاهشناپذیر هست یا نه صرف این که تشخیص دهیم حذف کدام بخشها دستیابی به عملکرد پایه از طریق سایر بخشها را غیرممکن میکند، کافی نیست. برای اطمینان، تشخیص این بخشهای غیرقابل کنار گذاشتن، گام مهمی برای تعیین پیچیدگیکاهشناپذیر در عمل است اما کافی نیست. به علاوه، لازم است ثابت کنیم که با هیچ سیستم سادهتری به همان عملکرد پایه دست نمییابیم. برای مثال، یک چهارپایه را در نظر بگیرید. فرض کنید که عملکرد پایهی این چهارپایه این است که بستری دارای ارتفاعی بیشتر از سطح زمین برای نشستن فراهم کند. از این نظر، هر کدام از پایهها برای دست یافتن به این عملکرد پایه غیرقابل حذفاند. (در صورتی که هر کدام از پایهها را حذف کنیم، دیگر نمیتوان با سایر بخشهای باقی مانده به عملکرد پایه دست یافت). با وجود این، از آنجا که میتوان با یک سیستم بسیار سادهتر به همین عملکرد پایه دست یافت (برای مثال با یک سنگ بلوک)، چهارپایه را نمیتوان یک مجموعهی کاهشناپذیر قلمداد کرد.
برای تعیین این که یک سیستم پیچیده کاهشناپذیر هست یا نه از دو راهکار میتوان استفاده کرد: (1) تحلیلی تجربی از سیستم با حذف بخشها (به صورت تکی یا گروهی) و سپس بازآرایی و انطباق سیستم با قسمتهای باقی مانده جهت تعیین این که آیا عملکرد پایهای سیستم میتواند با همین قسمتهای باقی مانده حاصل شود یا خیر. (2) تحلیل مفهومی از سیستم و به طور خاص از آن بخشهایی که حذف آنها بازیابی عملکرد پایه را ناممکن میسازد. این تحلیل برای این است که نشان دهد هیچ سیستمی متشکل از بخشهای کمتر نمیتواند عملکرد پایه را به اجرا درآورد. بخشهای غیرقابل حذف در بخش (1) مشخص میشوند و سپس در بخش (2) تایید میشود که هیچ سادهسازیای به هستهی کاهشناپذیر یک سیستم کاهشناپذیر تعلق ندارد. توجه داشته باشید که مرحله (1) و (2) میتوانند به صورت جداگانه یا یکجا اجرا شوند و اگر یکجا اجرا شوند نیاز به رعایت ترتیب خاصی نیست. بنابراین، ممکن است یک نفر ابتدا تحلیل نظری را انجام دهد تا تعیین کند که چه بخشهایی برای ارایهی یک عملکرد پایه ضروریاند و سپس به صورت تجربی اثبات کند که چه بخشهایی به طور قطع برای دستیابی به عملکرد پایه غیرقابل حذفاند. برای مثال، در مورد موتور قایقی که پیشتر صحبت کردیم، یک تحلیل نظری نشان میدهد که نمیتوان سیستمی داشت که عملکرد پایه را به اجرا در آورد اما پروانه، موتور یا میل گاردان نداشته باشد. در نتیجه این بخشها متعلق به هستهی کاهشناپذیرند، حقیقتی که میتواند سپس به صورت تجربی هم به اثبات برسد، بدین صورت که این بخشها را حذف کنیم و نشان دهیم که سایر قسمتهای باقی مانده توان بازیابی عملکرد پایه را ندارند.
پیچیدگیکاهشناپذیر تفاوتهایی آشکار با فرم دیگر پیچیدگی که «پیچیدگی تجمعی»[13] خوانده میشود، دارد. یک سیستم زمانی یک مجموعهی تجمعی است که بخشهای مختلف سیستم بتوانند به صورت پیدرپی بازآرایی شوند به طوری که حذف متوالی قسمتهای مختلف سیستم، هرگز منجر به نقص کامل عملکرد سیستم نشود. «شهر» مثالی از یک سیستم پیچیدهی تجمعی است. میتوانیم به طور مرتب افراد و خدمات را از شهر حذف کنیم تا جایی که تنها یک دهکدهی کوچک باقی بماند – بدون این که حتی لحظهای مفهوم جامعه را که در این حالت عملکرد پایهی شهر است، از دست بدهد. اگر حذف متوالی شهروندان و خدمات شهری را به صورت یک فیلم ضبط شده روی یک نوار فرض کنیم، آن وقت با معکوس کردن روند فیلم میتوانیم تکامل تدریجی شهر را مشاهده کنیم. تجمع تدریجی پیچیدگی طی فرگشت داروینی فرایندی رو به جلو است، به طوری که حرکت معکوس آن حذف متوالی اجزای تشکیل دهنده از یک سیستم پیچیدهی تجمعی است و البته این در حالی است که در هر گام از فرایند حذف، عملکرد سیستم همچنان حفظ میشود. این فرایند نشان میدهد که مکانیسم انتخاب داروینی را میتوان به راحتی به عنوان نوعی پیچیدگی تجمعی به حساب آورد.
اما پیچیدگیکاهشناپذیر چهطور؟ آیا مکانیسم انتخاب داروینی میتواند یک پیچیدگیکاهشناپذیر هم باشد؟ اگر انتخاب با توجه به یک هدف انجام شود، در این صورت این انتخاب برای تولید یک پیچیدگیکاهشناپذیر کار سختی در پیش ندارد. همان ساعت جیبی که پیشتر در موردش صحبت کردیم را در نظر بگیرید. اگر هدف ساخت یک گاهشمار دارای عملکرد باشد، آن گاه یک نفر میتواند یک فرایند انتخاب هدفمند را تعیین کند و طی آن یک فنر، یک صفحه، یک عقربهی ساعتشمار و یک عقربهی دقیقهشمار و سایر بخشهای غیرقابل حذف لازم برای ساخت یک ساعت جیبی را برگزیند و در نهایت همهی این بخشها را در کنار هم قرار دهد تا یک ساعت دارای عملکرد بسازد. به طور مشابه، میشود تصور کرد که یک موجود زنده برای ساخت یک ساختار جدید، طی چندین نسل با فراهم آوردن اجزای مشخص (شاید با تنوع تصادفی)، کنار هم قرار دادن آنها (با یک فرایند انتخاب هدفمند) و سپس زمانی که همهی اجزا کنار هم گردآمدند، با سر هم کردن آنها، یک ساختار جدید بسازد. با داشتن یک هدف از پیش معین، انتخاب کار دشواری برای ساخت یک سیستم پیچیدهی تقلیل ناپذیر در پیش نخواهد داشت.
با این شیوهی استدلال، انتخاب در زیستشناسی با یک مشکل بدیهی مواجه است. انتخابی که در زیستشناسی فعالیت میکند، انتخابطبیعی داروین است که این شکل از انتخاب بدون هیچ هدف، نقشه یا مقصد معینی کار میکند. انتخابطبیعی به هیچ آیندهای نگاه نمیکند، بلکه صرفا چشمش به حال است و میپرسد چه چیزی برای «اکنون» این ارگانیسم مفید است نه آینده یا چند نسل بعدی آن. انتخابطبیعی تنها به لذتهای آنی علاقهمند است، نه لذتهایی که با تاخیر بدست میآیند. او یک فرصت طلب است نه یک استراتژیست. این خصوصیات انتخابطبیعی در وهلهی اول محدودیت به نظر میآیند، اما برای زیستشناسانی که تمایل دارند پدیدهها را به صورت مطلقا فیزیکی و ماشینی نگاه کنند، این موضوع جذابیت خاصی دارد زیرا آنها ترجیح میدهند ظهور پیچیدگیهای زیستی را نتیجهی فرایند مادی غیرمستقیم و بنابراین جدای از طراحی در نظر بگیرند. با این حال وقتی داروین انتخاب را یک فرایند غیرمستقیم در نظر گرفت، بیدلیل انواع پیچیدگیهایی را که سیستمهای زیستی میتوانستند به اجرا درآورند، محدود کرد. با توجه به نظریهی داروین، سیستمهای زیستی به راحتی میتوانند پیچیدگی تجمعی را نشان دهند اما برای پیچیدگیکاهشناپذیر این نظریه راه دشواری در پیش دارد.
چرا این گونه است؟ مشکل این است که یک سیستم پیچیدهی کاهشناپذیر تنها زمانی به عملکرد پایهاش دست مییابد که تمام اجزای هستهی کاهشناپذیرش به طور همزمان در جایشان قرار گرفته باشند. در نتیجه، اگر انتخابطبیعی بخواهد عملکردی را برای یک سیستم پیچیدهی کاهشناپذیر انتخاب کند، یا باید تمام اجزای هستهی کاهشناپذیر را یکجا درست کند، یا اصلا نباید کاری کند. اگر سیستمهای مورد بحث ما سیستمهایی ساده بودند، آنگاه این موضوع مشکل خاصی محسوب نمیشد. اما این چنین نیست. سیستمهای پیچیدهی کاهشناپذیر زیستشیمیاییای که مایکل بیهی[14] در کتاب جعبه سیاه داروین[15] مطرح میکند، ماشینهای پروتئینی هستند که از پروتئینهای متعددی که هر کدام برای عملکرد پایهای ماشین ضروری و غیرقابل چشمپوشیاند، تشکیل شدهاند.
داروینیسم، به یک فرایند تکاملی تدریجی که طی آن عملکردها و فرایندها به صورت تدریجی شکل میگیرند متعهد است اما این موضوع امروز با یک معضل روبهرو است. فرگشت داروینی نمیتواند با عمل انتخابطبیعی و بهبود بخشیدن یک پیشساز سادهتر که از قبل عملکردی را نشان میدهد، سیستم پیچیدهی کاهشناپذیری بسازد که یک عملکرد پایهای خاص را به نمایش میگذارد. مشکل این است که این عملکرد خاص مورد نظر از قبل وجود ندارد و بنابراین انتخابطبیعی چیزی برای انتخاب کردن ندارد، مگر این که سیستم پیچیدهی کاهشناپذیر از قبل وجود داشته باشد. این نشان میدهد که فرگشت داروینی تنها زمانی میتواند یک سیستم پیچیدهی کاهشناپذیر دارای عملکرد پایه بسازد که سیستم از پیش موجود را که عملکرد متفاوتی ارایه میدهد بردارد و بازآرایی کند و سیستم پیچیدهی کاهشناپذیر مورد نظر را بسازد. اما همانطور که در ادامهی این مقاله خواهیم دید، هیچ شواهدی وجود ندارد که نشان دهد بازآراییهایی که برای شکلدادن به چنین سیستمهای پیچیدهی کاهشناپذیری مورد نیازند اصولا رخ دادهاند، چه برسد به این که فرایند فرگشت تدریجی داروین آنها را به طور مناسب هماهنگ کرده باشد. در عوض، شواهد نشان میدهند که هر گونه بازآرایی به چنان هماهنگی عظیمی در سیستم در حال بازآرایی احتیاج دارد که حصول سیستم پیچیدهی کاهشناپذیر را به طور کامل دور از دسترس فرگشت داروینی قرار میدهد. البته چنین هماهنگی عظیمی نشان از طراحی دارد.
داروین در منشا انواع تاکید میکند که نظریهاش یک نظریهی تدریجی است به طوری که طی آن ساختارهای پیچیدهی زیستی (به تعبیر او «اندامهای پیچیده») باید ظرفیت شکلگیری را طی آنچه که او «تغییرات متعدد، پی در پی و جزیی» میخواند داشته باشند]4[. این نکته نشان میدهد که نظریهی داروین تا حدی تضمین میکند که زیستشناسان میتوانند مسیرهای دقیق و قابلآزمایش داروینی را ترسیم کنند تا از طریق آنها ساختارهای بیولوژیکی پیچیده از طریق تغییرات متعدد، پیدرپی و جزیی ایجاد شوند. متقابلا، این نظریه تا این حد متزلزل است که زیستشناسان نه تنها نمیتوانند چنین مسیرهایی را ارایه دهند، بلکه همچنین دلایل روشنی هم دارند که نشان دهد چنین مسیرهایی اصولا وجود ندارند. بدین ترتیب، پیچیدگیکاهشناپذیر ماشینهای پروتئینی با قدرت نظریهی داروین را رد میکند. علاوه بر این، از آنجا که پیچیدگیکاهشناپذیر در سطوح زیستشیمیایی اتفاق میافتد، هیچ سطح بنیادیتری برای تحلیلهای زیستی وجود ندارد که بتوان پیچیدگیکاهشناپذیر ماشینهای پروتئینی را به آنها ارجاع داد و کور سوی امیدی به موفقیت تحلیلهای داروینی از منظر انتخابطبیعی و تنوع تصادفی داشت. زیستشیمی زیربنایی حیات همان شیمی و فیزیک معمولی است و هیچکدام هم نمیتوانند پیچیدگی زیستی را توضیح دهند.
یک ماشین پروتئینی پیچیده که مشخصا توجه بسیاری از محافل زیستشناسی را به خود جلب کرده است، تاژک باکتری است. در یک سخنرانی عمومی، هوارد برگ[16]، زیستشناس دانشگاه هاروارد، تاژک باکتری را «کارآمدترین ماشین جهان» نامیده است. تاژک یک موتور چرخشی است که نیروی آن را اسید تامین میکند و یک دم شلاق مانند دارد که با چرخش خود باکتری را در محیط آبی به جلو میراند. این دم شلاق مانند، شبیه یک پروانه عمل میکند که با دهها هزار دور در دقیقه میچرخد و میتواند جهت خود را با زاویهی نود درجه تغییر دهد. ساختار پیچیدهی تاژک شامل یک موتور چرخان، یک پایه، یک حلقهی O، غلافها، صفحهی نگهدارنده، میلهی گردان، یک پیشران، یک قلاب برای پیوستن به این پیشران و یک موتور با پیشرانهی اسیدی است.
عملکرد پایهی تاژک باکتری این است که با چرخاندن سریع دو سویهی دم شلاق مانندش، باکتری را در محیط آبی به پیش براند. توجه داشته باشید که یک دم شلاق مانند با این خصوصیات زمانی که به عنوان ساختار حرکتی جستوجو برای غذا مورد استفاده قرار میگیرد، دیگر یک عضو لوکس و تجملی محسوب نمیشود، بلکه عضوی ضروری و حیاتی برای باکتری به شمار میرود. برای پیش بردن یک باکتری در محیط آبی، تاژک باید بر حرکت براونی (حرکت تصادفی مولکولهای آب که مدام به اجسام کوچک معلق در آب تنه میزند) غلبه کند. لازم است که تاژک در هر دو جهت بچرخد زیرا حرکت براونی باکتری را از مسیرش در راستای منبع تغذیه منحرف میکنند. معکوس شدن جهت چرخش دم باعث میشود باکتری چرخی بزند و بار دیگر خودش را در مسیر غذا قرار دهد و تلاش کند که به آن دست یابد. در تاژکی که میخواهد یک باکتری را در یک محیط آبی در جهت منبع غذایی به حرکت درآورد، عملکرد حداقلی باید این باشد که بتواند دم شلاق مانندش را در هر دو جهت و بینهایت سریع به چرخش در آورد. تاژک باکتری بالاتر از 10000 دور دقیقه میچرخد (در حقیقت، نزدیک به 20000 دور و حتی 100000 دور در دقیقه). اساسا هر مقداری کمتر از این مانع غلبهی باکتری بر اثر منحرف کنندهی حرکت براونی میشود و بدین ترتیب جلو رسیدن باکتری به غلظت مناسبی از مواد معدنی مورد نیاز برای زنده ماندن، تولید مثل و رشد را میگیرد]5[.
ساختمان پیچیدهی تاژک به برهمکنش هماهنگ حدود سی پروتئین نیاز دارد که بیستواندی پروتئین دیگر هم در فرایند گرد آمدن آنها کنار یکدیگر دخیلاند. با این حال فقدان هر کدام از این پروتئینها منجر به از دست رفتن عملکرد موتور میشود ]6[. این پروتئینها هستهی کاهشناپذیر تاژک را میسازند. این هسته چهقدر پیچیده است؟ جان پستگیت[17] بخشی از این پیچیدگی را شرح میدهد:
ما اکنون میدانیم که یک تاژک معمولی باکتری، یک رشتهی لولهمانند بلند پروتئینی است. این رشته به طور شُل و آزادی پیچ خورده است، مانند یک فنر کشیده شده یا شاید یک در باز کن بطری و در انتهایش هم به دیوارهی سلولی نزدیک میشود و یک ناحیهی ضخیم و انعطافپذیر میسازد. از آنجا که انتهای این رشته اغلب به صورت خمیده است، قلاب نامیده میشود. دیوارهی سلولی یک پوستهی نسبتا سفت دارد که درون آن اندکی نرمتر است و در مرکز آن یک پروتوپلاسم ژلهمانند قرار دارد. تاژک و قلاب آن به این (یا درون این) پوسته متصل میشود و یک ویژگی بسیار قابل توجه شیوهای است که این دو بخش به همدیگر متصل میشوند. در باکتریای به نام باسیلوس سوبتیلیس، قلاب مانند یک میله از میان دیوارهی خارجی امتداد مییابد و در چند نانومتر انتهایی این میله دو دیسک قرار دارد. یکی از آنها تقریبا به طور کامل در انتهای تاژک قرار دارد، به طوری که گویی در غشای داخلی است، همان غشایی که پروتوپلاسم سلول را میپوشاند و دیسک نزدیک انتها تقربیا در دیوارهی سلولی قرار میگیرد. در عمل، این تاژک بلند به نظر میرسد که با قلابش در جای خودش نگه داشته شده است، به طوری که دو دیسک مانند دو مهره یا یک مهره و یک واشر عمل میکنند]7[.
این نقل قول تنها شمهای از پیچیدگی تاژک باکتری را نشان میدهد. در اینجا پستگیت توضیح میدهد که پروانهی پیشبرندهی باکتری چهطور ساخته میشود و چهطور به دیوارهی سلولی متصل میشود. به علاوه، نیاز به موتوری هم هست که بتواند این پروانه را به حرکت در آورد. این موتور باید بر پایهای سوار شود و بر سر جایش ثابت شود. همچنین باید قادر باشد که در هر دو جهت بچرخد. پیچیدگیها به سرعت افزایش مییابند و یک تحلیل نظری نشان میدهد که تاژک باکتری یک هستهی کاهشناپذیر بینهایت پیچیده را به منصه ظهور مینشاند.
خوب، پس منشا تاژک باکتری چیست؟ از دیدگاه داروینی، تاژک طی مکانیسم انتخاب داروینی و از یک باکتری که نه تنها فاقد تاژک بوده بلکه حتی تمام ژنهای کدکنندهی پروتئین تاژکی را هم نداشته (از جمله هر ژنی که هومولوگ ژنهای تاژک باشد) شکل گرفته است. برای این که با مکانیسم داروینی بتوان یک تاژک باکتریایی ساخت، تغییرات تصادفی ژنتیکی باید ژنهایی را که این پروتئینها را کد میکنند، بوجود بیاورد. سپس انتخابطبیعی باید این پروتئینها را حفظ کند و همگی آنها را در یک باکتری گرد آورد و به شیوهای صحیح آنها را سرهم کند. چهطور چنین چیزی امکانپذیر است؟ در ادامهی این مقاله به بررسی میزان غیرممکن بودن این تبیین داروینی میپردازیم و خواهیم دید که طراحیهوشمندانهانه تبیین بسیار متقاعدکنندهتری در این زمینه ارایه میدهد.
۲- استدلالی از دل پیچیدگی کاهشناپذیر
در جعبه سیاه داروین مایکل بیهی ایدهی پیچیدگیکاهشناپذیر را مطرح میکند و سپس استدلال میکند که پیچیدگیکاهشناپذیر ماشینهای پروتئینی شواهد متقاعد کنندهای بر وجود یک طراح حقیقی در زیستشناسی دارد. از زمان انتشار این اثر در سال 1996، کتاب بیهی به طور گسترده در بسیاری از نشریههای مشهور و مجلات علمی مورد بررسی قرار گرفته است]۸[. این اثر همچنین به طور گسترده در اینترنت به بحث گذاشته شده است]9[. بسیاری از منتقدین اعتراف کردهاند که بیهی حقایق علمیاش را کاملا سرراست ارایه داده است. آنها همچنین ادعا کردهاند که جای یک توضیح دقیق نوداروینی در مورد نحوهی پیدایش ماشینهای پروتئینی پیچیدهی کاهشناپذیر در مباحث زیستشناسی خالی است. با این حال آنها نقدهای نظری و روششناختیای نیز به استدلال وی وارد کردهاند. بیهی چیری را ارایه میدهد که میتوان آن را «استدلالی مبتنی بر پیچیدگیکاهشناپذیر»[18] نامید. این استدلال مدعی است که پیچیدگیکاهشناپذیر سیستمهای زیستی فراتر از دسترسی مکانیسم فرگشت تکاملی است و این که تنها طراحی است که میتواند توضیحی مناسب برای آنها ارایه دهد.
چهگونه استدلال از پیچیدگیکاهشناپذیر به چنین نتیجهای میرسد؟ متاسفانه نقدها به این استدلال به دو شیوه وارد میشود و هیچ کدام هم منصفانه نیستند. این نقدها به طور معمول تمایل دارند این استدلال را یا به صورت کاملا منطقی یا به شکل کاملا تجربی در نظر بگیرند. دیدگاه منطقی چنین است: ساختارهای معینی وجود دارد که میتوان ثابت کرد با مکانیسم داروینی غیرقابل دسترساند. این ساختارها از خصوصیتIC (برای مثال، پیچیدگی کاهشناپذیر) برخوردارند. اما ساختارهای زیستی معینی هم وجود دارند که آنها هم از IC برخوردارند، در نتیجه آنها باید با مکانیسم داروینی غیرقابل دسترس باشند. نقطه نظر تجربی چنین است: ساختارهای زیستی خاصی هستند که به طرز وحشتناکی پیچیدهاند. حتی یک توضیح ساده هم در متون زیستی نیست که بگوید مکانیسم داروینی چهگونه ممکن است آنها را ساخته باشد. پس احتمالا چیزهایی فراتر از انتخابطبیعی مسئول منشا آنها بوده است.
بنابر مطالبی که بیان شد، این دو دیدگاه تفاوتهایی بنیادین با همدیگر دارند و به توجیهات متفاوتی هم احتیاج دارند. اگر استدلال حاصل از پیچیدگیکاهشناپذیر یک دیدگاه منطقی محض ارایه میکرد، در این صورت باید مثل قانون دوم ترمودینامیک (که با عنوان نظریهی ارگودیک[19] شناخته میشود) که برای جلوگیری از حرکت دایمی ماشینها مورد استفاده قرار میگیرد، به طور کامل از ریاضیات پیروی میکرد. اما اگر این استدلال یک نقطه نظر تجربی محض باشد، آن گاه به نظر میرسد که چیزی بیش از یک استدلال ناشی از غفلت نیست و صرفا نشان میدهد که مسیرهای فرگشتیای که به برخی از سیستمهای زیستی خاص منتهی میشوند هنوز نیاز به توضیح دارند، حقیقتی که منتقدان به راحتی آن را میپذیرند.
با توجه به دیدگاه منتقدان، نه دیدگاه منطقی و نه تفسیر تجربی و نه حتی ترکیبی از آن دو چالشی پیش پای نظریهی فرگشت نمیگذارند. بیایید این دو را بررسی کنیم. از دید منطقی، پیچیدگیکاهشناپذیر هرگز نمیتواند تمام راهها را به روی فرگشت داروینی ببندد. پیچیدگیکاهشناپذیر تضمین میکند که هیچ کدام از بخشهای هستهی کاهشناپذیر سیستم قابل حذف نیستند. از این نظر که اگر هر کدام از قسمتهای هسته را حذف کنید، آنگاه دیگر نمیتوانید عملکرد پایهای سیستم را از قسمتهای باقی مانده باز یابید. اما این موضوع امکان حذف قسمتها یا جدا سازی بخشهایی که عملکردهای پایهای دیگری را انجام میدهند باز میگذارد. (عملکردی که به احتمال زیاد میتواند موضوعی برای فشار انتخاب باشد). پیچیدگی کاهشناپذیر، که به عنوان یک محدودیت منطقی مطلق به آن نگریسته میشود، آنگاه یک روزنهی گریز برای مکانیسم داروینی به جا میگذارد. به طور خاص، این امکان را باز میگذارد که مسیرهای ناشناختهی غیرمستقیم داروین بتوانند یک سیستم پیچیدهی زیستی را با سیستمهای دیگری که عملکردهای متفاوتی نسبت به سیستم مورد نظر دارند، تکامل بخشد.
دید تجربی اما به نظر میرسد صرفا مرتکب مغالطهی متعارف استدلال از غفلت شده است. گیریم که سیستمهای زیستی خاصی به طرز اعجابانگیزی پیچیدهاند و ما هم نمیدانیم که چهگونه نشات یافتهاند، حالا که چه؟ این بدان معنا نیست که مکانیسم داروینی یا برخی مکانیسمهای مادی دیگر آن را انجام ندادهاند بلکه صرفا ممکن است به این معنا باشد که ما فعلا نمیدانیم که آن مکانیسمها چهگونه این کار را کردهاند. ترکیب دیدگاههای منطقی و تجربی اما بیاعتبارترین گزینه است زیرا باعث میشود هواداران طراحیهوشمندانهانه محکوم به دو پهلوگویی و تقلب شوند به طوری که انگار آنها از پیچیدگیکاهشناپذیر استفاده میکنند تا هر جا که نیاز بود دیدگاه تجربی یا منطقی را برحسب مصحلت ارایه دهند.
این تکذیبیهها برای رد استدلال بیهی زیادی کوتهنظرانه و آساناند. در حقیقت استدلالی که بیهی از دل پیچیدگیکاهشناپذیر ارایه میدهد بسیار ظریفتر و دقیقتر از هر کدام از پیشنهادهایی است که منتقدان مطرح میکنند. این استدلال را به طور کامل میتوان در سه دیدگاه مطرح کرد: دیدگاه منطقی، تجربی و تبیینی. بسیار بیشتر از آن که این دیدگاهها همدیگر را نقض کنند، با همدیگر کار میکنند و یکدیگر را تقویت میکنند. دیدگاه منطقی چنین است: میتوان اثبات کرد که ساختارهای خاصی هستند که به طور «مستقیم» با روش داروینی غیرقابل دسترسیاند. آنها به طرز خاصی پیچیدهی کاهشناپذیرند. اما ساختارهای زیستی معینی هم وجود دارند که آنها هم پیچیدگیهای کاهشناپذیر خاصی دارند. پس آنها هم باید با مسیرهای داروینی غیرقابل دسترس باشند. این صورتبندی به نظر مشابه دیدگاه قبلی به نظر میرسد اما از یک جنبهی بسیار مهم متفاوت است. در قاعدهی پیشین، عدم دسترسی نسبت به مکانیسم داروینی در نظر گرفته شده بود و هیچ محدودیتی هم برای آن نبود. بنابراین، تمام مسیرهای داروینی را در برمیگرفت چه مستقیم و چه غیرمستقیم. اما در اینجا ما تنها مکانیسمهای داروینیای را در نظر میگیریم که دربردارندهی مسیرهای مستقیم داروینی هستند.
یک مسیر داروینی مستقیم، مسیری است که یک سیستم با انتخابطبیعی و ارتقای تدریجی یک عملکرد خاص، تکامل مییابد. همچنان که سیستم تکامل مییابد، عملکرد تکامل نمییابد بلکه در جای خود ثابت میماند. بنابراین، ممکن است تصور کنیم که در تکامل قلب، عملکرد آن از ابتدا این بوده که خون را پمپ کند. در این مورد یک مسیر داروینی شاید آن را توجیه کند. از سوی دیگر، ممکن است تصور کنیم که در تکامل قلب عملکرد اولیهی آن این بوده که صدایی مهیب تولید کند تا شکارچیان را از موجود دور کند و بعدها به عنوان پمپ کنندهی خون مورد استفاده قرار گرفته است. در این مورد یک مسیر داروینی غیرمستقیم برای توضیح دادن آن مورد نیاز خواهد بود. در اینجا مسیر غیرمستقیم است، زیرا نه تنها سیستم تکامل یافته بلکه عملکرد سیستم هم تکامل یافته است. حال، از منظر دیدگاه منطقی، استدلال ناشی از پیچیدگیکاهشناپذیر تنها با مسیرهای مستقیم داروینی سر و کار دارد. این از خود تعریف پیچیدگیکاهشناپذیر مشهود است، زیرا هستهی کاهشناپذیر را دقیقا در ارتباط با یک عملکرد منفرد به نام عملکرد پایهای سیستم پیچیدهی کاهشناپذیر تعریف کردیم. (عملکردی که بدون همهی قسمتهای هستهی کاهشناپذیر نمیتوانست وجود داشته باشد).
در رد مسیرهای مستقیم داروینی منتهی به سیستمهای پیچیدهی کاهشناپذیر، استدلال ناشی از پیچیدگیکاهشناپذیر میگوید که سیستمهای زیستشیمیایی پیچیدهی کاهشناپذیر کاملا دور از دسترس مسیرهای مستقیم داروینی قرار دارند. چهطور میتوانیم بفهمیم که چنین سیستمهایی واقعا دور از دسترس مسیرهای داروینی هستند یا نه؟ آنچه را که من به عنوان سیستم پیچیدهی کاهشناپذیری تعربف کردم که قرار است با یک مسیر مستقیم داروینی تکامل یابد، در نظر بگیرید. در این حالت سیستم باید از تکامل سیستمهای سادهتری که همان عملکرد را نشان میدهند، نشات گرفته باشد. اما از آنجا که هستهی کاهشناپذیر یک سیستم پیچیدهی کاهشناپذیر را نمیتوان بدون تخریب عملکرد پایه ساده کرد، آن وقت دیگر هیچ پیشینه تکاملیای با هستهی سادهتر که بتواند همان عملکرد را ارایه دهد، نمیتواند وجود داشته باشد. در نتیجه، تنها راهی که مسیر مستقیم داروینی میتواند یک سیستم پیچیدهی کاهشناپذیر را تکامل دهد این است که آن را به طور یکباره و با یک اتفاق خیلی نامحتمل یا شانسی تکامل داده باشد. نتیجه این است که نسبت دادن پیچیدگیکاهشناپذیر به مسیر داروینی مانند این است که ساخته شدن چهرههای روی کوه راشمور را به باد و فرسایش نسبت دهیم. یک احتمالی وجود دارد که باد و فرسایش کوه راشمور را کندهکاری کرده باشد، اما چندان واقعگرایانه نیست.
اثبات غیرقابل دسترس بودن سیستمهای پیچیدهی کاهشناپذیر برای مسیرهای مستقیم داروینی عملی احتمالاتی است. این اثبات اگر چه از منطق و ریاضیات بهره میبرد، اما مسیرهای مستقیم داروینی را به عنوان پدیدهای کاملا غیرممکن رد نمیکند. از نظر منطقی ممکن است هر چیزی بتواند با رخدادهایی تصادفی و حتی خیلی غیرمحتمل به هر چیز دیگری تبدیل شود. برای مثال، از نظر منطقی محتمل است که یک بازیکن آماتور شطرنج مجموعهای حرکات بینظیر از خود نشان دهد و آن وقت یک استاد بزرگ شطرنج را در مسابقه شکست دهد. اما رخ دادن چنین اتفاقی مغایر محدودیت توانمندی بازیکن آماتور در بازی شطرنج خواهد بود نه به دلیل آن. به طور مشابه، اگر یک مسیر مستقیم داروینی به یک سیستم پیچیدهی زیستشیمیایی دست یابد، این اتفاق مغایر خصوصیات ذاتی یا ظرفیتهای مکانیسم داروینی است نه به دلیل آن. بنابراین، وقتی گفته میشود که سیستمهای زیستشیمیایی پییچدهی کاهشناپذیر با مسیرهای مستقیم داروینی غیرقابل دسترساند، منظور هواداران طراحی این است که مکانیسمهای داروینی ظرفیت ذاتی برای ساخت چنین سیستمهایی ندارند مگر با اتفاقاتی بسیار نامحتمل و تصادفی.
به هر حال، منتقدان استدلال ناشی از پیچیدگیکاهشناپذیر تلاش نمیکنند تا فهرستی از مسیرهای مستقیم داروینی ارایه دهند که به سیستمهای پیچیدهی کاهشناپذیر ختم شده است، بلکه سعی میکنند این کار را با مسیرهای غیرمستقیم داروینی انجام دهند. در مسیرهای غیرمستقیم داروینی، یک سیستم با حفظ و ارتقای عملکرد موجود تکامل نمییابد بلکه با تغییر مداوم آن عملکرد تکامل مییابد، در حالی که در مسیرهای مستقیم ساختارها تکامل مییابند اما عملکرد ثابت باقی میماند. به عبارت دیگر، در مسیر غیرمستقیم هم ساختار و هم عملکرد هر دو تکامل مییابد. این اثر متقابل تکامل به هم پیوستهی ساختار و عملکرد با عنوان «تکامل همراه»[20] شناخته میشود.
اما استدلال ناشی از پیچیدگیکاهشناپذیر چهگونه با مسیر غیرمستقیم داروین سازگار میشود؟ این موضوع با دیدگاه منطقی بررسی نمیشود بلکه از منظر دیدگاه تجربی به آن نگاه میشود. حقیقت این است که برای سیستمهای زیستشیمیایی پیچیدهی کاهشناپذیر هیچ مسیر غیرمستقیم داروینی شناخته نشده است. در بهترین حالت، زیستشناسان قادر بودند زیرمجموعههایی از چنین سیستمهایی جدا کنند که سایر عملکردها را به نمایش میگذارند. اما هر ماشینی که به طرزی منطقی پیچیده باشد، شامل زیرمجموعههایی است که عملکردی مجزا از ماشین اصلی انجام میدهند. پس صرف وقوع یا شناسایی زیرسیستمهایی که خودشان بتوانند عملکردی متفاوت ارایه دهند، دلیلی بر این نیست که مسیرهای غیرمستقیم داروینی میتوانند به چنین سیستمهایی منتهی شوند. آنچه مورد نیاز است، یک تبیین یکپارچهی داروینی است که توضیحی مفصل و قابل آزمودن ارایه دهد؛ توضیحی که نشان دهد چهطور زیرسیستمهایی که تحت تکامل همراه قرار میگیرند میتوانند به صورت تدریجی به یک سیستم پیچیدهی کاهشناپذیر تبدیل شوند. هیچ تبیینی از این دست وجود ندارد یا تا حالا ارایه نشده است. در واقع، اگر چنین تبیینی وجود میداشت منتقدان طراحیهوشمندانهانه تنها کافی بود که به آنها ارجاع دهند و طراحیهوشمندانه را به طور کامل رد کنند.
از این نظر حرکت معمول منتقدان طراحیهوشمندانه این است که دست بالا را میگیرند و استدلال ناشی از پیچیدگیکاهشناپذیر را متهم میکنند که استدلالی ناشی از بیاطلاعی است. راه معمول برای صورتبندی این نقد این است که میگویند «فقدان شواهد، شاهدی بر فقدان نیست.» اما مانند بسیاری از عبارات قدیمی این هم نیاز به بررسی دقیق دارد. مطمئنا این حکم در مورد بسیاری از شرایط و موقعیتهای روزمره صدق میکند. تصور کنید کسی با تمام وجود همهی خانه را برای پیداکردن دسته کلید ماشینش میگردد؛ زیر هر وسیله را جستوجو میکند، از افراد دیگر به عنوان نیروی کمکی درخواست میکند و صبح روز بعد، در حالی که دیگر هیچ امیدی به یافتنش ندارد، آن را بیرون خانه روی ماشینش پیدا میکند. در این مورد نبود شواهد، پیش از پیدا کردن دستهکلید، به این معنا نبود که کلیدی وجود ندارد. با این حال اطلاع از وجود داشتن کلیدها در همان آغاز شاهدی بر حضورشان بود.
اما اگر مطمئن نبودیم که کلیدها اصولا وجود دارند یا نه، چه؟ وضعیت زیستشناسی تکاملی از این هم بغرنجتر است. شاید کسی مطمئن نباشد که دسته کلید فرضی ما وجود دارد، اما حداقل میتواند اطمینان حاصل کند که کلید ماشین به طورکلی وجود دارد. مسیرهای غیرمستقیم داروینی که پیچیدگیکاهشناپذیر را توجیه میکنند بیشتر شبیه این است که فرض کنیم یک لپرکان[21] (نوعی موجود خیالی در اساطیر سنتی ایرلندی) در اتاق کودک پنهان شده است. از آنجا که شواهد دال بر وجود نداشتن لپرکانها تمام و کمال است، دیگر خیلی غیرمنطقی است که بگوییم «فقدان شواهد، شاهدی بر فقدان نیست.» و بر همین مبنا حضور لپرکان را جدی بگیریم. این کار اساسا همان جستوجوی کاملا بیفایدهی مدافعان نظریهی تکامل برای یافتن مسیرهای داروینی غیرمستقیم معتبر برای تبیین پیچیدگیکاهشناپذیر است.
اگر بعد از بارها تلاش و جستوجوی همهی مکانهای احتمالی، چیزی را که دنبالش میگشتید، نیافتید و هیچ مدرکی هم وجود نداشته باشد که پی ببرید آن چیزی که به دنبالش میگردید آیا اصولا در واقعیت وجود دارد یا نه، آن وقت خیلی منطقی است که با خودتان فکر کنید شاید آن چیزی که به دنبالش میگردید اصلا وجود خارجی نداشته باشد. این استدلالی از دیدگاه پیچیدگیکاهشناپذیر در مورد مسیرهای غیرمستقیم داروینی است. قضیه صرفا این نیست که ما چنین مسیرهایی را در مورد تاژک باکتری (ماشین زیستشیمیایی پیچیدهی کاهشناپذیری که نماد محافل طراحیهوشمندانه است) نمیدانیم، موضوع این است که ما هیچ مسیری برای چنین سیستمهایی نمیشناسیم. در اینجا فقدان کاملا «سیستماتیک و فراگیر» است. به این دلیل است که برخی منتقدان داروینیسم مانند فرانکلین هرولد[22] و جیمز شاپیرو[23] (که هیچ کدامشان هوادار طراحیهوشمندانه نیستند) میگویند این که فرض کنیم مسیرهای غیرمستقیم داروینی برای چنین سیستمهایی فعلا کشف نشده است، «آرزویی خیالپردازانه» است]10[.
به طور خلاصه، استدلال ناشی از پیچیدگیکاهشناپذیر به یک دیدگاه منطقی و یک دیدگاهی تجربی میانجامد. از دیدگاه منطقی، پیچیدگیکاهشناپذیر ساختارهای زیستی ورای دسترسی مسیرهای مستقیم داروینی قرار دارد. از دیدگاه تجربی، ناکامی زیستشناسی تکاملی در کشف مسیرهای غیرمستقیم داروینی که به ساختارهای پیچیدهی کاهشناپذیر منتهی میشوند، کاملا فراگیر و سیستماتیک و بنابراین شواهدی است برای شک یا حتی رد مسیرهای غیرمستقیم داروینی در پاسخ دادن به پیچیدگی کاهشناپذیر. دیدگاههای منطقی و تجربی با هم کیفرخواستی ویرانگر علیه مکانیسم داروینی شکل میدهند. مکانیسمی که به طور معمول فرض میشود قابلیت حل همهی مشکلات پیچیدگیهای زیستیای را که در اشکال مختلف حیات ظاهر میشوند، دارد. با وجود همهی اینها، دیدگاههای منطقی و تجربی پاسخی در این باره نمیدهند که چهطور میتوان از ناکامی داروینیسم در توضیح سیستمهای پیچیدهی کاهشناپذیر به مشروعیت استفاده از طراحی در توجیه آنها رسید.
این جا جایی است که استدلال ناشی از پیچیدگی کاهشناپذیر نیاز دارد تا دیدگاه سومی به نام دیدگاه تبیینی ارایه دهد. تبیینهای علمی به اشکال و ظواهر گوناگونی نمایان میشوند، اما یک چیز هست که هیچ تبیین علمی نمیتواند فاقد آن باشند و آن «کفایت علیتی»[24] است. یک توضیح علمی لازم است که قدرت علیتی کافی برای توضیح مفهوم مورد پرسش را داشته باشد. در غیر این صورت این مفهوم بدون توضیح باقی میماند. در این جا مفهومی که مورد پرسش قرار گرفته پیچیدگیکاهشناپذیر مجموعهای از ماشینهای زیستشیمیایی خاص است. چنین سیستمهایی چهگونه بوجود آمدهاند؟ این اتفاق از مسیرهای مستقیم داروینی بوقوع نپیوسته است. پیچیدگیکاهشناپذیر این را با دلایل منطقی و ریاضیاتی رد میکند. همچنین با مسیرهای غیرمستقیم داروینی هم اتفاق نیفتاده است. در اینجا نبود شواهد علمی به اندازهی نبود شواهد در مورد لپرکانها کامل است. توسل به سازوکارهای ناشناختهی مادی هم در این زمینه کمکی نمیکند، زیرا در این خصوص نه تنها شواهد مورد نیاز به طور کامل وجود ندارند، بلکه نظریهای که برایش هیچ مدرکی وجود ندارد هم در حقیقت وجود خارجی ندارد.
بنابراین، وقتی بحث در مورد سیستمهای زیستشیمیایی پیچیدهی کاهشناپذیر باشد، هیچ شواهدی وجود ندارد که بپذیریم سازوکارهای مادی برای به وجود آوردن آنها کفایت علّی دارند. اما در مورد آگاهی و هوش چهطور؟ اینها برای ساخت سیستمهای پیچیدهی کاهشناپذیر بسیار مشهورند (برای مثال انسان به طور مدوام ماشینهایی میسازد که پیچیدگیکاهشناپذیر را به نمایش میگذارد). بدین ترتیب هوش کفایت لازم برای ساخت پیچیدگیکاهشناپذیر را دارد. بنابراین، دیدگاه تبیینی استدلال ناشی از پیچیدگیکاهشناپذیر این است که براساس اصل کفایت علیتی، طراحیهوشمندانه توضیح علمیتری برای پیچیدگیکاهشناپذیر سیستمهای زیستشیمیایی نسبت به مکانیسم داروینی است.
برای ایجاد دیدگاههای منطقی و تجربی، استدلال ناشی از پیچیدگیکاهشناپذیر با تشخیص محدودیتهای مکانیسم داروینی نقشی نقادانه یا منفی را به عهده میگیرد. در مقابل، در دیدگاه تبیینی اما این استدلال نقشی مثبت و سازنده دارد و زمینهی مثبتی را فراهم میکند تا چنین بیندیشیم که پیچیدگیکاهشناپذیر سیستمهای زیستشیمیایی در حقیقت طراحی شدهاند. یک پرسش در مورد این دیدگاه همچنان باقی است. دیدگاه منطقی مسیرهای مستقیم داروینی منتهی به پیچدگی کاهشناپذیر را رد میکند و دیدگاه تجربی این کار را در مود مسیرهای غیرمستقیم داروینی انجام میدهد. اما همان طور که هیچ شواهد تجربی برای مسیرهای مستقیم داروینی که به پیچیدگیکاهشناپذیر منتهی میشوند وجود ندارد، این موضوع در مورد مسیرهای غیرمستقیم نیز صدق میکند. بنابراین، از آنجا که دیدگاه تجربی میتواند هر دو مسیر داروینی را حذف کند و کنار بزند، ممکن است به نظر بیاید دیدگاه منطقی اضافی است. اما چنین نیست و درحقیقت دیدگاه تجربی هرگز نمیتواند به اندازهی دیدگاه منطقی پرونده را علیه داروینیسم سنگین کند.
اگر به بهترین نمونههای تایید شدهی فرگشت داروینی در متون زیستی (از زمان داروین تاکنون) نگاه کنید، خواهید دید که انتخابطبیعی به طور پیوسته خصوصیت معینی که عملکردی خاص را به شیوهای معین به اجرا در میآورد، بهبود میبخشد. در واقع خود مفهوم «بهبود بخشیدن» (که مفهومی کلیدی در منشا انواع داروین است) دلالت بر این دارد که یک چیز معین و از پیش موجود دارد از یک جنبهی خاص بهتر میشود. بهبود یافتن از این نظر مطابق مسیر مستقیم داروینی است. در مقابل، یک مسیر غیرمستقیم داروین (که طی آن یک عملکرد خاص جایش را به عملکرد دیگری میدهد، بنابراین دیگر نمیتواند بهبود پیدا کند چون دیگر وجود ندارد) گرچه اغلب به کمک دادههای فسیلی و مولکولی از سوی زیستشناسان استنباط میشود، اما تایید دقیق و قطعی آنها کار بسیار دشوارتری است.
فهمیدن دلیلش هم چندان سخت نیست؛ طبق تعریف، انتخاب طبییعی عملکردهای موجود را انتخاب میکند. به عبارت دیگر، عملکردهایی که در حال حاضر وجود دارند و به طرقی به موجود زنده کمک میکنند. از سوی دیگر، انتخابطبیعی نمیتواند عملکردهای آینده را انتخاب کند. عملکردهایی که وجود ندارند و به طرقی در حال حاضر به موجود زنده کمک میکنند، برای انتخابطبیعی نامرییاند. اولین بار که عملکردی پا به عرصهی وجود میگذارد، مکانیسم داروینی میتواند عمل انتخاب را روی آن اعمال کند. اما گذار از یک عملکرد قدیمی به عملکردی جدید، کاری نیست که مکانیسم داروینی از پس آن برآید. سیستمی که عملکردی از قبل موجود و قابل گزینش را به نمایش میگذارد، چهگونه میتواند به یک سیستم جدید تبدیل شود که عملکرد قابل گزینش جدیدی را ارایه میدهد؟ از آنجا که انتخابطبیعی تنها میتواند عملکردهای موجود را انتخاب کند، در اینجا کمکی از دستش بر نمیآید و تمام کار بر دوش دگرگونی تصادفی است تا در آن زمان حساس که عملکردها تغییر میکنند، با اصلاحات درست و مورد نیاز وارد شود (یا همانطور که داروین از آن یاد میکند، «تا زمانی که دگرگونیهای ثمربخش وجود دارند، انتخابطبیعی هیچ کاری نمیتواند بکند»]11[.) با این حال، هیچ شواهد حقیقی و قطعیای وجود ندارد که نشان دهد دگرگونی تصادفی میتواند اصلاحات متوالی مورد نیاز برای تکامل پیچیدگیکاهشناپذیر را ایجاد کند.
استدلال ناشی از پیچیدگی کاهشناپذیر، با بیان این دیدگاه منطقی که پیچیدگیکاهشناپذیر مسیرهای مستقیم داروینی را رد میکند، شکلی از فرگشت داروینی را که به بهترین نحو تایید شدهاند رد میکند. در مقابل، مسیرهای غیرمستقیم داروینی، آن قدر متغیر و دگرگونیپذیرند که نمیتوان آنها را به صورت علمی مورد آزمون قرار داد، مگر این که با دقت زیاد مشخص شوند. وقتی موضوع در مورد سیستمهای پیچیدهی کاهشناپذیر باشد، همواره این مسیرها کاملا غیرمشخص و نامتعیناند به طوری که نمیتوان آنها را تایید یا رد کرد. با این دیدگاه، استدلال ناشی از پیچیدگیکاهشناپذیر تا جایی که میتواند در محدود کردن مکانیسم داروینی پیش میرود و همهی ملاحظات تجربی لازم را در نظر میگیرد تا هیچ روزنهی گریزی باقی نماند. از آنجا که استنباطهای منطقی به طور ذاتی قویتر از استنباطهای تجربیاند، استدلال ناشی از پیچیدگیکاهشناپذیر در رد مکانیسم داروینی، تا حد امکان محکم و قوی است. مسئله فقط این نیست که مجموعهای از سیستمهای زیستی خاص آن قدر پیچیدهاند که نمیتوانیم تصور کنیم با مکانیسمهای داروینی تکامل یافته باشند، بلکه میتوانیم به طور قاطع نشان دهیم که مسیرهای مستقیم داروینی برای ساخت چنین سیستمهایی عملا فاقد صلاحیتاند و مسیرهای غیرمستقیم هم (که همواره اثباتشان دشوارتر بوده) به طور کامل فاقد پشتوانهی تجربی کافی برای چنین کاری هستند. در مقابل ما میدانیم که چه چیزی قدرت کافی برای ساخت پیچیدگیکاهشناپذیر را دارد: طراحیهوشمندانه.
3- معماری و قوسهای رومی
حالا که تعاریف و منطق زیربنایی استدلال ناشی از پیچیدگیکاهشناپذیر معلوم شد، بیاید دو استدلال اصلی مخالف را که داروینیستها علیه آن مطرح کردهاند، بررسی کنیم. من در این بخش به یکی از این استدلالها میپردازم و در بخش بعد به دیگری. این استدلالهای مخالف تلاش میکنند نشان دهند که اگر از نزدیک بررسی کنیم میبینیم که یک سیستم پیچیدهی کاهشناپذیر از طریق تغییرات تدریجی و بدون طراحی ساخته میشود. با توجه به استدلال داربستی، برای این که تکامل بتواند یک سیستم پیچیدهی کاهشناپذیر بسازد، ابتدا نیاز است که چند سیستم پیچیدهی کاهشپذیر با جهش ساخته شود و انتخاب، اجزایی را به آنها اضافه کند. سپس در مقاطعی، یک زیرسیستم ظهور پیدا میکند که عملکردی مستقل و خودکار (فارغ از سایر اجزای سیستم) دارد. از آنجا که این زیرسیستم میتواند به طور خودکار کار کند، سایر بخشها زاید خواهند بود و به دور انداخته میشوند. زمانی که همهی این بخشهای اضافی به دور انداخته شدند، یک سیستم پیچیدهی کاهشناپذیر خواهیم داشت. به طور خلاصه، آن چه که به نظر یک تفاوت کیفی است در حقیقت تنها نتیجهی تعداد زیادی تغییرات کوچک کمّی است.
بنابراین استدلال داربستی ادعا میکند که حذف اضافات عملکردی راهی محتمل برای رسیدن به پیچیدگیکاهشناپذیر است. بنابراین به جای آن که تکامل با فرایندی پایین به بالا و افزودن تدریجی بخشهای مختلف به یک سیستم، به پیچیدگیکاهشناپذیر دست یابد، فرض میشود که پیچیدگیکاهشناپذیر از فرایندی بالا به پایین و حذف اجزای اضافی یک سیستم حاصل میشود. برای نمونه، وضعیتهایی وجود دارد که مطابق آنچه توماس اشنایدر[25] میگوید «یک گونهی دارای عملکرد بدون یک سیستم کنترل ژنتیک خاص میتواند زنده بماند، اما اگر چنین کنترلی از ابتدا وجود داشته باشد، بهتر این کار را انجام میدهد»]12[. اشنایدر در ادامه اضافه میکند:
هر عملکرد جدید باید چنین خصوصیتی داشته باشد تا حیاتِ گونه به آن وابسته شود، به طوری که گویی یکی از راههای اولیهی بقا است که اگر نباشد موجود در اثر ضعف بدنی از بین میرود. من چنین وضعیتی را «قوس رومی» میخوانم، چرا که بعد از این که چنین ساختاری روی یک داربست ساخته میشود، میتوان داربست را حذف کرد. در سیستمهای زیستی نیز چنین است که این داربستها زمانی که دیگر مورد نیاز نباشند حذف میشوند. این قوسهای رومی در زیستشناسی بسیار رایجاند و یکی از پیامدهای طبیعی فرایندهای تکاملی محسوب میشوند.]13[.
برای ساخت یک قوس رومی به یک داربست احتیاج است. تا زمانی که این داربست در جای خودش است، قطعات کمان میتوانند از جایگاه خودشان بیرون بیایند و جابهجا شوند. اما به محض این که تمام قطعات در جای خودشان قرار گرفتند و داربست حذف شد (مانند حذف شدن بخشهای زاید) هر کدام از قطعات قوس تبدیل به بخشی غیرقابل حذف میشود و کمان هم یک سیستم پیچیدهی کاهشناپذیر را شکل میدهد.
اما در اینجا دو مشکل وجود دارد. اولا اگر بخواهیم دقیق و با کمی سختگیری نگاه کنیم میبینیم که قوس رومی پیچیدهی کاهشناپذیر نیست. بله هر کدام از قطعات تشکیل دهندهی قوس برای بودنش الزامیاند و اگر هر کدامشان را حذف کنید دیگر نمیتوانید با بازآرایی سایر قسمتها یک کمان بسازید. اما یک قوس رومی قابل سادهسازی است. یک قطعه سنگ را میتوان تراشید و به شکل همان کمان در آورد و میبینیم که همان عملکرد را به همان روش و سبک و سیاق انجام میدهد. با وجود این، ممکن است کسی بگوید حالا اگر قوس رومی با کمی سختگیری پیچیدهی کاهشناپذیر نباشد، چندان هم مهم نیست. چنین قوسی با همهی این حرفها بالاخره از نظر عملکردی یکپارچه است و بنابراین پرسشی که باقی میماند این است که آیا داربستها میتوانند به طورکلی راه قابل قبولی برای رسیدن به یک سیستم یکپارچهی دارای کارکرد، و به طور خاص، به یک سیستم پیچیدهی کاهشناپذیر ارایه دهند؟
با وجود این، یک مشکل جدیتر در مورد استدلال داربستی وجود دارد. فرض کنید اگر قرار بود فرگشت داروینی با داربست یک سیستم پیچیدهی کاهشناپذیر مثل تاژک باکتری درست کند، آن وقت این موضوع دقیقا چه معنایی میداشت. مکانیسم انتخاب داروین بر اساس بهرهگیری از مزیتها یا انتخاب یک عملکرد موجود کار میکند. علاوه بر این، بدیهی است که یک سیستم پیچیدهی کاهشناپذیر مانند تاژک باکتری عملکرد پایهای قابل انتخاب داشته باشد. در نتیجه، تاژک به علاوهی هر داربستی که فرض کنیم، همان عملکرد پایه را نشان میدهد، بنابراین داربست که حالا دیگر اضافی است میتواند با انتخابطبیعی حذف شود. حالا اجازه دهید این پرسش را طرح کنیم که هنگام ساخت یک سیستم یکپارچه از یک سیستم پیچیدهی کاهشناپذیر به علاوهی داربست، دقیقا چه زمانی عملکرد پایه شکل گرفت؟ برای مثال، در مجموعهی تاژک باکتری و داربست، حرکت چرخش دو سویه برای پیش بردن باکتری در محیط آبی دقیقا چه زمانی شکل گرفت؟
داربست زدن هیچ تاثیری در این حقیقت ندارد که عملکرد پایهای یک سیستم کاهشناپذیر، بنا به تعریف، تنها زمانی شکل میگیرد که تمام اجزای سیستم جای خودشان باشند. با فرض این که داروینیسم بتواند توضیحی مبتنی بر داربست برای سیستم پیچیدهی کاهشناپذیر ارایه دهد، چالش داروینیست این است که بتواند توالیای از میانجیهای «عملکردی» تدریجی را که منجر به ساخت چنین سیستمی میشوند، شناسایی کند. برای این کار او باید از چند سیستم اولیهی ساده شروع کند و در نهایت به یک سیستم پیچدهی کاهشناپذیر به همراه یک داربست برسد و بعد نشان دهد که انتخابطبیعی داربست را زمانی که تبدیل به بخشی زاید شده است، حذف میکند. گرچه داربست میتواند به ساخت یک سیستم پیچیدهی کاهشناپذیر کمک کند، داربست به طور خاص منطبق با عملکرد پایهای سیستمی است که دارد به ساختش کمک میکند (برای مثال تاژک). علاوه براین، تنها شاهد دال بر آن عملکرد پایه از خود سیستم پیچیدهی کاهشناپذیر بدست میآید. بنابراین، برای این که مکانیسم داروینی بتواند از طریق داربستزنی یک سیستم پیچدهی کاهشناپذیر بسازد، سیستم و داربست باید عملکردی «متفاوت» داشته باشند تا زمانی که تمام اجزای هستهی سیستم پیچیدهی کاهشناپذیر نهایی فراهم شوند، در جای خود قرار بگیرند و یک سیستم دارای عملکرد را شکل دهند. اما در این صورت دیگر اصطلاح داربست مناسب نخواهد بود. وقتی از داربست صحبت میکنیم منظور ساختاری است که عملکردی معین ارایه میدهد نه این که یک ساختار در حال تکامل باشد که عملکردش هم تکامل یابد. این ما را به استدلال مخالف بعدی میرساند که داروینیستها علیه استدلال ناشی از پیچیدگیکاهشناپذیر اقامه میکنند.
4- تکامل همراه و همکاری
برای توضیح پیچیدگی کاهشناپذیر، داروینیستها همواره در نهایت دست به دامان مسیرهای غیرمستقیم داروینی میشوند. در یک مسیر غیرمستقیم داروینی نه تنها ساختار تکامل مییابد بلکه عملکرد آن هم توامان تکامل مییابد. در مقابل، در یک مسیر مستقیم، انتخابطبیعی ساختاری را ارتقا یا بهبود میدهد که از پیش عملکردی را از خود نشان میدهد و در این میان خود عملکرد تغییری نمیکند. از آنجا که یک سیستم پیچیدهی کاهشناپذیر تا زمانی که هستهی کاهشناپذیرش به طور کامل سر جایش قرار نگرفته باشد، عملکردی از خودش نشان نمیدهد، اگر یک مسیر مستقیم داروینی بخواهد چنین سیستمی بسازد باید آن را به صورت یکباره نازل کند. اما این مهملی بیش نیست است. این سیستمها به طرز شگفتانگیزی پیچیدهاند، اگر قرار باشد که بدون هر گونه طراحی ساخته شده باشند، باید همان طور که داروین میگوید از طریق «اصلاحات متعدد، متوالی و جزیی» شکل گرفته باشند]14[. بنابراین، تنها راهی که داروینیسم میتواند پیچیدگیکاهشناپذیر را توضیح دهد این است که بگوید این ساختارها از طریق مسیرهای غیرمستقیم داروینی شکل گرفتهاند که طی آن ساختارها و عملکردها همراه با یکدیگر تکامل مییابند.
یک راه برای آن که چنین چیزی بتواند اتفاق بیفتد این است که بخشهایی که قبلا برای اهداف متفاوتی در نظر گرفته شده بودند از هم جدا شوند و در سیستم جدید به خدمت گرفته شوند. درست مثل قطعات خودرو، دوچرخه، قایق موتوری و قطار که ترکیب میشوند و بازآرایی میشوند تا بالاخره یک هواپیما را میسازند. نظریهپردازهای تکاملی گاهی از آنها با عنوان «چهلتکه»[26] یاد میکنند. بنابراین همهی هواپیماها چهلتکههایی از مصالح از پیش موجودی هستند که در اصل کاربردهای دیگری برایشان در نظر گرفته شده بود. هیچ غیرممکن منطقیای وجود ندارد که بخواهد از ساخت سیستمهای پیچیدهی کاهشناپذیر به کمک این چهلتکهها جلوگیری کند. اما یک چهل تکه، اگر به طور خیلی دقیق و عالی کار کند، آن وقت نیاز است که توضیحی در مورد نحوه شکلگیریاش وجود داشته باشد. برای مثال، تاژک باکتری یک معجزهی مهندسی در عملکرد و کوچکسازی است. این که چنین سیستمی را صرفا چهلتکهای بخوانیم که از وصله و پینه شدن مصالح از پیش موجود ساخته شده است، چندان روشنگرانه نیست و پاسخی هم در مورد نحوهی شکلگیری و منشا آن ارایه نمیدهد.
مشکل تلاش برای توضیح دادن یک سیستم پیچیدهی کاهشناپذیر مثل تاژک باکتری به عنوان یک چهل تکه این است که نیازمند چندین همکاری همزمان است. قضیه صرفا این نیست که یک چیز برای عملکردی تکامل مییابد و سپس، شاید بدون هیچ تغییری، برای عملکردی کاملا متفاوتی مورد استفاده قرار گیرد (مثلا سنگی را فرض کنید که قبلا روی کاغذها قرار میدادند تا باد آنها را نبرد اما حالا به عنوان پادری استفاده میشود تا جلو بستهشدن دری را بگیرد). مشکل این است که چندین بخش پروتئینی از سیستمهای عملکردی مختلف باید جدا شوند سپس همگی به طور یکپارچه در هم ادغام شوند تا یک سیستم یکپارچهی جدید را بسازند (همان طور که هواپیما از بخشهای مختلف خودرو، دوچرخه، قایق و قطار ساخته شده است).
حتی اگر همهی این قسمتهای مورد نیاز (برای مثال پروتئینها) برای ساخت یک تاژک باکتری درون سلول حضور داشته باشند اما عملکردهای متفاوتی انجام دهند، هیچ دلیلی وجود ندارد که فکرکنیم این بخشها میتوانند به طور خودبهخودی کنار هم جمع شوند و یک سیستم به شدت یکپارچه مانند تاژک را بسازند. در اینجا مشکل این است که بخشهایی که در سیستمهای مجزا دارای عملکردند خیلی بعید است که به گونهای با هم منطبق شوند که بتوانند به طور منسجم با یکدیگر در یک سیستم واحد کار کنند. یک پیچ را تصور کنید که بخشی از یک سیستم است و یک مهره که بخشی از یک سیستم دیگر است. اگر این سیستمها منشاهایی مستقل از هم داشته باشند، مانند بسیاری از سیستمهای تکاملیافتهی زیستی، خیلی بعید است که پیچ و مهره به هم بخورند و از نظر مکانیکی مفید باشند (مثلا مهره نه خیلی تنگ باشد که کلا پیچ درونش نرود، نه خیلی گشاد باشد که دیگر پیچ با آن درگیر نشود).
این مشکل درون سلول حتی بزرگتر است. تکامل تاژک باکتری را در نظر بگیرید؛ علاوه بر پروتئینهایی که درون تاژک قرار میگیرند، سلولی که تاژک درون آن تکامل مییابد، کلی پروتئین دیگر دارد که هیچ نقش قابل توجهی در تاژک ندارند. در واقع بخش اعظم پروتئینهای درون سلول به این صورتاند. با این شرایط، چهطور ممکن است فقط و فقط همان پروتئینهایی که درون تاژک عملکرد دارند گرد هم بیایند و به محل درستشان هدایت شوند بدون این که با سایر پروتئینها برهمکنش ایجاد کنند؟ مثل این است که وارد یک هایپرمارکت بشویم و با چشمان بسته و به صورت تصادفی چیزهایی از قفسهها برداریم و بعد انتظار داشته باشیم که در نهایت آنچه در چرخ خریدمان جمع کردهایم دقیقا چیزهای مورد نیاز برای پخت یک کیک باشد. چنین نتیجهای به شدت نامحتمل است. آلن اور[27]، زیستشناس دانشگاه روچستر[28]، که از هواداران طراحیهوشمندانه نیست در این باره میگوید:
ممکن است فکر کنیم که بعضی از بخشهای یک سیستم پیچیدهی کاهشناپذیر برای اهداف خاصی به صورت گام به گام تکامل یافتهاند و سپس یکباره برای عملکردی جدید به خدمت گرفته شدهاند. اما این خیلی نامحتمل است. مثل این است که انتظار داشته باشید جعبه دندهی ماشینتان خیلی ناگهانی به کیسهی هوا کمک کند. چنین اتفاقاتی ممکن است خیلیخیلی به ندرت رخ دهند اما مطمئنا راه حل کلیای برای پیچیدگیکاهشناپذیر ارایه نمیدهند]1۵[.
مشکل این نوع سناریوهای همکاری این است که نیاز به چندین همکاری همزمان از چندین سیستم دارای عملکرد دارند تا یک سیستم پیچیدهی کاهشناپذیر را شکل بدهد.
اما اگر به جای آن این همکاری خیلی تدریجیتر و تجمعیتر اتفاق بیفتد چه؟ تصور کنید که در تکامل تاژک باکتری، انتخابطبیعی به صورت تدریجی بخشهای پروتئینی از پیش موجود را در یک ساختار واحد به همکاری وادارد و ساختاری بسازد که عملکردش همراه با ساختارش تکمیل میشود. در این صورت یک سیستم پیچیدهی کاهشناپذیر شکل میگیرد که از اجزایی ساخته شده که قبلا قابل حذف بودند اما حالا تبدیل به اجزایی غیرقابل حذف تبدیل شدهاند (مانند بخشهایی که برای ساخت هستهی یک سیستم پیچیدهی کاهشناپذیر مورد نیازند). آلن اور این موضوع را این گونه به تصویر میکشد:
یک سیستم پیچیدهی کاهشناپذیر میتواند به صورت تدریجی با اضافهکردن بخشهایی ساخته شود که ابتدا صرفا مزیتهایی داشتند اما بعد – به دلیل تغییراتی که در ادامه متحمل میشود – تبدیل به بخشی حیاتی برای سیستم میشود. منطق پشت این امر بسیار ساده است. برخی قسمتها (A) ابتدا کاری انجام میدهد (شاید هم آن را خیلی خوب انجام ندهد). بخشی دیگر (B) بعدا اضافه میشود تا به A کمک کند. این بخش جدید بخش ضروریای نیست اما صرفا باعث بهبود چیزها میشود. اما در ادامه A (یا چیزی دیگر) ممکن است تغییری کند که باعث بشود تبدیل به جزیی غیرقابل حذف شود. این فرایند، همچنان که بخشهای دیگری در سیستم ادغام میشود، ادامه مییابد و در نهایت قسمتهای متعددی وجود خواهند داشت که برای عملکرد سیستم حیاتی محسوب میشوند]16[.
بیاید این استدلال را ارزیابی کنیم. اور یک افزایش تدریجی در پیچیدگی را به تصویر میکشد که در آن بخشهای جدیدی که عملکرد را ارتقا میدهند متناوبا به صورت بخشهایی غیرقابل حذف در میآیند. اما در مورد کدام عملکرد داریم حرف میزنیم؟ بدیهی است که عملکردهای میانهی مسیر باید با عملکرد نهایی متفاوت باشند زیرا عملکرد نهایی از سوی یک سیستم پیچیدهی کاهشناپذیر ارایه میشود و از این رو هیچ سیستمی که اساسا هستهی کاهشناپذیر سادهتری دارد نمیتواند آن عملکرد را ارایه دهد. اما بیاید برویم سراغ مشکل تجربی: هیچ شواهد تجربیای وجود ندارد که نشان دهد سیستمهای زیستشیمیایی پیچیدهی کاهشناپذیری مانند تاژک باکتری به این روش و با اضافه شدن اجزایی که بعدا تبدیل به اجزایی جداییناپذیر از سیستم شدهاند شکل گرفته باشد.
در واقع اور همراستا با سایر اعضای جامعهی داروینی، هیچ وقت چیزی بیش از یک سناریوی به شدت انتزاعی در مورد نحوهی بروز پیچیدگیکاهشناپذیر ارایه نمیدهند. واضح است که چیزی بیش از این مورد نیاز است. حداقل چیزی که مورد نیاز است، یک دستورالعمل یا مدل دقیق و قابل آزمودن است که نشان دهد مسیرهای غیرمستقیم داروینی چهگونه ممکن است ماشینهای پیچیدهی کاهشناپذیر زیستشیمیایی مانند تاژک باکتری را شکل داده باشند. اور در عوض این کار را با سیستمهای انتزاعی نامعین و به صورت شماتیکی و با حروفی مانند A و B انجام میدهد. زیستشناسان تکاملی هیچ چیزی مانند یک مسیر تکاملی دقیق که به سیستمهای پیچیدهی کاهشناپذیری مانند تاژک باکتری منتهی شود، ندارند.
نزدیکترین چیزی که زیستشناسان توانستهاند به عنوان یک پیشساز تکاملی احتمالی برای تاژک باکتری شناسایی کنند، با عنوان سیستم ترشحی نوع III یا همان (TTSS) شناخته میشود. TTSS نوعی پمپ است که باکتریهای بیماریزای خاصی را قادر میسازد پروتئینهایی که عامل بیماریزایی هستند (پروتئینهای ویرولانت) را به درون موجودات میزبان تزریق کنند. یکی از باکتریهایی که TTSS را در اختیار دارد «یرسینیا پستیس»[29] است؛ ارگانیسمی که مسئول طاعون سیاهی است که در قرن چهاردهم یک سوم جمعیت اروپا را به کام مرگ فرستاد. TTSS همان سیستم توزیع و پخشی بود که یرسینیا پستیس به وسیلهی آن باعث تباهی گستردهی حیات انسانها شد. اکنون مشخص شده است که حدود ده تا از پروتئینهایی که در ساخت TTSS دخیلاند مشابه (هومولوگ) پروتئینهایی هستند که در تاژک باکتری یافت میشوند. به علاوه، TTSS تقریبا با بخشی از تاژک که در ساخت رشتهاش مورد استفاده قرار گرفته است، مطابقت دارد (برای مثال دم شلاق مانند). اما توجه داشته باشید که این چنین هم نیست که بتوانیم TTSS را با آن بخش مربوطه در تاژک جایگزین کنیم و تاژکمان همچنان عملکرد داشته باشد، چون پروتئینهای درون TTSS با پروتئینهای تاژک باکتری منطبق نیستند و بعد از جایگزینی عملکرد نخواهند داشت.
علی رغم همهی این دشواریها برای مرتبط دانستن TTSS به تاژک باکتری، بیایید فرض کنیم که TTSS زیرمجموعهای از تاژک است. اگر چنین باشد، عملکردی مجزا از تاژک را نشان میدهد. با همهی اینها یافتن زیرسیستمی از یک سیستم که دارای عملکرد متفاوتی باشد به سختی میتواند مبنای استدلالی قرار گیرد که براساس آن نتیجه بگیریم سیستم اصلی از آن زیرسیستم تکامل یافته است. کسی ممکن است بگوید از آنجا که موتور یک موتورسیکلت خودش میتواند به عنوان یک وسیلهی گرمکننده عمل کند، بنابراین موتورسیکلت از موتور تکامل یافته است. شاید هم همین طور باشد، اما نه بدون طراحیهوشمندانه. در واقع، تقریبا همهی سیستمهایی که از چندین بخش به شدت یکپارچه تشکیل شدهاند، بخشهای متعددی دارند که میتوانند عملکردهای متفاوتی انجام دهند. در بهترین حالت TTSS یک گام احتمالی در مسیر غیرمستقیم تکامل داروینی در تکامل تاژک باکتری است. اما این همچنان راهحلی برای تکامل تاژک باکتری ارایه نمیدهد. آنچه ما احتیاج داریم یک مسیر تکاملی کامل است، نه یک آبادی احتمالی در میانهی مسیر. ادعایی غیر از این مثل این است که بگوییم چون جزایر هاوایی را کشف کردهایم پس میتوانیم با پای پیاده از لسآنجلس تا توکیو برویم.
یک مشکل دیگر هم وجود دارد. هدف از مطرح کردن TTSS به طور کلی ارایهی آن به عنوان پیشساز تکاملی تاژک باکتری بود. با وجود این، بهترین شواهدی که زیستشناسان تکاملی در حال حاضر به آن دست یافتهاند نشان میدهد که TTSS از تاژک باکتری تکامل یافته باشد نه بالعکس]17[. به شکلی شهودی درک این موضوع آسان است که TTSSمحتملتر است از تاژک باکتری تکامل یافته باشد نه بالعکس. تاژک باکتری یک ساختار حرکتی است که باکتری را در محیط آبی به پیش میراند. آب از زمان پیدایش حیات در محیط وجود داشته است. در واقع زیستشناسان تکاملی تخمین میزنند که عمر تاژک باکتری حدود 2 تا 3 میلیارد سال باشد اما TTSS یک سیستم تحویل و توزیع برای باکتریهای بیماریزا در گیاهان و جانوران است. بنابراین، عملکرد آن وابسته به وجود موجود زندهی پرسلول است. با این تفاسیر، TTSS تنها میتواند بعد از پیدایش موجودات پرسلول، یعنی تقریبا از 600 میلیون سال پیش، وجود داشته باشد.
در نتیجه، TTSS توضیحی در مورد تکامل تاژک ارایه نمیدهد. در بهترین حالت این تاژک باکتری است که میتواند تکامل TTSS را توضیح دهد. اما حتی این هم کاملا درست نیست. با همهی اینها TTSS بسیار سادهتر از تاژک است. TTSS تقریبا شامل ده پروتئین است که هومولوگ پروتئینهای تاژکاند. تاژک به حدود سی یا چهل پروتئین بیشتر نیاز دارد که هر کدامشان به نوبهی خود منحصر به فردند. تکامل باید ظهور پیچیدگی از سادگی را توضیح بدهد اما اگر TTSS از تاژک تکامل یافته باشد، آن وقت تمام کاری که کردهایم این است که یک چیز سادهتر را بر مبنای یک چیز پیچیدهتر توضیح دادهایم.
علی رغم این مشکلات، داروینیستها همچنان اصرار به ارایه TTSS به عنوان پیشساز تکاملی تاژک باکتری دارند]1۸[. برخی از آنها حتی از این هم فراتر میروند و ساختارهای بینابینیای معرفی میکنند که طی آنها TTSS به تاژک باکتری تکامل یافته است]19[. اما این ساختارهای بینابینی به عنوان پیشسازهای تاژک باکتری، حتی از خود TTSS هم متزلزلترند. برخلاف TTSS، این حدواسطها تنها در ذهن زیستشناسان تکاملی وجود دارند و نه در طبیعت و نه در آزمایشگاه نمیتوان آنها را پیدا کرد و زیستشناسان تکاملی هم هیچ وقت با جزییات کامل آنها را توصیف نمیکنند که بشود آنها را شناسایی کرد تا ببینیم آیا واقعا وجود دارند یا نه. داروینیستها چنین حدواسطهایی را پیشنهاد میکنند و ادعا میکنند که گامهای بینابینیای فراهم کردهاند که طی آنها TTSS به تاژک باکتری منتهی میشود. برخی حتی ادعا میکنند که با فراهم کردن این حدواسطهای خیالی یک توجیه «مفصل، قابل راستی آزمایی و گام به گام» برای تشکیل تاژک باکتری ارایه دادهاند]20[. اما تمام اینها خیالاتی آرزوگونهاند.
برای ساختهشدن تاژک باکتری مطابق همین ایدهی بازسازی، باید گامهای زیر طی شود 1) یک باکتری دارای یک «TTSS اجدادی» را در نظر بگیرید تا چرخ تکاملی به راه بیفتد. 2) در مرحلهی بعد تصور کنید که این باکتری یک رشتهی مو مانندی (پیلوس)[30] را تکامل میدهد که به عنوان TTSS از باکتری بیرون میزند؛ این پیلوس بعدا با تکامل یافتن به تاژک تبدیل به «پروانهای» میشود که باکتری را در محیط آبی به پیش میبرد. 3) فرض کنید که این پیلوس در اثر انتخاب دستخوش مجموعهای از بهبودهای سریع مانند افزایش قدرت، کاهش شکستگیها، افزایش سرعت مونتاژ و غیره میشود. 4) سپس فرض کنید این پیلوس که در اصل جهت چسبیدن به سطوح تکامل یافته بود، حالا تحرکاتی را تکامل میدهد که ابتدا بسیار خام و ناشیانهاند و با پراکندگی تصادفی در جهات نامعلومی به وقوع میپیوندند. 5) سپس فرض کنید که در مرحلهی بعد این «پیشتاژک خام دارای عملکرد» چندین تحول جدید مانند کموتاکسی کسب میکند و بنابراین این حرکت جهتدار و در تعامل با محیط میشود. 6) در نهایت فرض کنید که معایب این سیستم کلی با انتخابطبیعی برطرف میشود و یک قلاب و سایر جزییات محوری اضافی هم به آن افزوده میشود و بدین ترتیب یک تاژک مدرن شکل میگیرد]21[.
برای توجیه چنین مدلی، داروینیستها باید نشان دهند که هر کدام از مراحل بالا به طور منطقی از مرحلهی قبل پیروی میکند. این مستلزم امکان ارزیابی احتمال گذار از یک مرحله به مرحله بعدی است و این به نوبهی خود متضمن این است که ساختارهای زیستی در هر مرحله با جزییات کامل توصیف شده باشند تا ارزیابی احتمال گذار بین مراحل مختلف امکانپذیر باشد. داروینیسم نظریهای است که به اتصال نقاط در یک پیکربندی فضایی زیستشناختی متکی است. اصل سخن این نظریه این است که شما میتوانید نقطهی A را به نقطهB در فضای پیکربندی زیستی متصل کنید به شرطی که بتوانید گامهایی به اندازهی کافی کوچک بردارید و در هر گام هم شایستگی زیستی افزایش یابد (یا حداقل شایستگی تغییری نکند). این گامها باید کوچک باشند چون داروینیسم نظریهای است که بر تغییرات تدریجی تجمعی مبتنی است به طوری که هر گام در طول مسیر به صورت منطقی امکانپذیر باشد. همانطور که داروین در کتاب منشا انواع اشاره میکند، برای این که نظریهاش به واقعیت بپیوندد باید بتواند پیچیدگیهای زیستی را به صورت «اصلاحات متعدد، متوالی و جزیی» توضیح دهد. هر توضیح دیگری باعث میشود که این نظریه به طور کامل نامحتمل شود.
آیا گذار از یک مرحله به مرحلهی بعد در مدل قبلی از نظر منطقی امکانپذیر است؟ آیا هر کدام از این مراحل، همان طور که داروین لازم میدانست، تنها از «اصلاحات جزیی» تشکیل شده است؟ هیچ راهی نیست که حتی بتوانیم پاسخگویی به این پرسشها را آغاز کنیم چون این مدل اصولا جزییات کافی ندارد. همهی زیستشناسان تکاملی در حقیقت هنوز هم پاسخی برای TTSS مدرن، تاژک باکتری مدرن و بسیاری از ساختارهای هومولوگی که در تاژکهای موجودات توسعهیافتهی امروزی دیده میشود، ندارند. زیستشناسان تکاملی نه حدواسطهایی را که این مدل نیاز دارد، در اختیار دارند و نه آن TTSS اجدادی را که این مدل با آن آغاز میکند. آنها حتی نمیدانند این حدواسطها چه شکلی هستند و دقیقا چه خصوصیات زیستشیمیاییای دارند. نمیدانند این سیستمهای بینابینیای که مدل مفروض میگیرد، اصولا کار میکنند یا نه. هیچ راهی نیست که بتوان تعیین کرد برای مکانیسم داروینی چهقدر آسان یا سخت است که بین گامهای مختلف در این مدل پل بزند. زیستشناسان تکاملی معمولا به مضاعف شدن ژنها و جهشها در نقاط کلیدی متوسل میشوند؛ جایی که فرض میشود مکانیسم داروینی به طور معقولی محتمل است که بر این گذارها تاثیر بگذارد. اما دقیقاچه ژنهایی مضاعف میشوند؟ و کدام ناحیه از چه ژنی صامت میشود؟
زیستشناسان تکاملی هیچ گاه حرفی در این باره نمیزنند. در واقع، گامها در این مدلها بسیار نامشخصاند و اگر به دنبال جزییات باشید، چنان مایوستان میکنند که این سوال عملا غیرقابل پاسخ باقی میماند. اما تا وقتی که پاسخهای دقیقی برای این پرسشها نداشته باشیم، هیچ راهی نیست که بدانیم گذارهایی که این مدلها توصیف میکنند از نظر منطقی محتملاند و همانهایی هستند که نظریهی داروین آنها را الزام میکند یا نه. در نتیجه، چنین مدلهایی غیرقابل راستیآزماییاند. برای آزمودن چنین مدلهایی نیاز است که بتوانیم در این مدل احتمال گذار از یک مرحله به مرحلهی بعد را ارزیابی کنیم. با وجود این، از آنجا که سیستمهای حدواسطی که در مراحل مختلف توصیف میشوند به طور کامل فاقد جزییاتاند (این سیستمها کاملا فرضیاند، تا جایی که میدانیم در طبیعت وجود ندارند، در آزمایشگاه در دسترس نیستند و محققان هیچ دستورالعملی برای ساخت آنها در آزمایشگاه ندارند)، مدلْ هیچ راهی برای چنین ارزیابیای پیش پای ما قرار نمیدهد.
بنابراین جای تعجب نیست که متون علمی در ارایهی پیشنهادی با جزییات کافی، قابل آزمودن و گام به گام از نحوهی تکامل همراهی که به ساخت سیستمهای زیستشیمیایی با پیچیدگیکاهشناپذیر منتهی میشود، دستشان خالی باشد. در خصوص چنین پیشنهادهایی، داروینیستها صرفا میگویند از آنجا که زیرسیستمهای سیستمهای پیچیدهی کاهشناپذیر میتوانند دارای عملکرد باشند، هر عملکرد این چنینیای میتواند از طریق انتخابطبیعی گزینش شود و با این نگرش دور از انتظار، سرخوشانه نتیجه میگیرند که انتخاب روی آن قسمتها کار میکند و سیستمهای پیچیدهی کاهشناپذیر میسازد]22[. اما این نتیجهگیری کاملا بیپایه و اساس است و همان طور که زیستشناس سلولی، فرانکلین هرولد[31] صراحتا بیان میکند، «در حال حاضر هیچ توجیه داروینی برای تکامل هیچکدام از سیستمهای زیستشیمیایی یا سلولی وجود ندارد مگر انواعی از گمانهزنیهای آرزوگونه.»]23[. لین مارگولیس[32]زیستشناس هم با همین صراحت میگوید: «مثل یک دونات شکری که موقتا اشتهایمان را ارضا میکند اما ما را از دریافت مواد مغذی بیشتر محروم میسازد، نوداروینیسم هم کنجکاوی ما را با انتزاعاتی که فاقد جزییات واقعیاند، ارضا میکند.»]24[.
به عنوان جمعبندی، مکانیسم داروینی اگر قرار باشد که کار کند نیازمند یک عملکرد قابل انتخاب است. به علاوه، مطابق مکانیسم داروینی قطعات عملکردی مختلفی که از سیستمهای مختلف با تکامل همراه گرد هم جمع شدهاند، از طریق مکانیسم داروینی قابل انتخاباند. اما آنچه در اینجا قابل انتخاب است عملکردهای منفرد از قطعات منفرد است و نه عملکردی از سیستمی که قرار است ساخته شود. مکانیسم داروینی عملکردهای از پیش موجود را انتخاب میکند و عملکردهایی را که قرار است در آینده از راه برسند، انتخاب نمیکند. به محض این که آن عملکرد شناسایی شد، مکانیسم داروینی میتواند انتخابش کند اما گذار از عملکرد فعلی به یک عملکرد جدید بخش سخت ماجرا است. چهگونه یک سیستم میتواند از یک قطعهی دارای عملکرد که در عملکرد خودشان قابل انتخاب شدناند به سیستمی برسد که از این قطعات استفاده میکند ولی عملکرد جدیدی را نمایان میسازد؟ مکانیسم داروینی در مورد ماشینهای زیستشیمیایی پیچیدهی کاهشناپذیر کمکی نمیکند.
5 – در رابطه با پیچیدگی مشخص
من در کتابهای استنباط طراحی[33] و ناهار مجانی در کار نیست[34]، معیاری برای شناسایی طراحی توصیف کردهام که پیچیدگی مشخص[35] نامیده میشود]25[. در این مقاله دیدیم که پیروان داروین هیچ توضیح مشروح، قابل آزمودن و گام به گامی در مورد تکاملِ هیچیک از ماشینهای زیستشیمیایی پیچیدهی کاهشناپذیر مانند تاژک باکتری ارایه نمیدهند. علاوه بر این، اگر پیشفرض متعصبانهی داروینیسم نتیجهگیریهایمان را رنگآمیزی نکند، ما به طور طبیعی تمایل داریم تا چنین سیستمهای پیچیدهی کاهشناپذیری را به عنوان محصولات طراحیهوشمندانه ببینیم. تمام شهود ما در این راستا عمل میکند. به همین دلیل است که ریچارد داوکینز[36] مینویسد «زیستشناسی علم مطالعهی مسایل پیچیدهای است که در ظاهر برای هدف خاصی طراحی شدهاند.» ]26[. همچنین به همین دلیل هم فرانسیس کریک[37] مینویسد، «زیستشناسان باید به طور مداوم این را در ذهن داشته باشند که آنچه میبینند طراحی نشده، بلکه تکامل یافته است.»]27[. با توجه به گفتههای داوکینز و کریک و سایر داروینیستهای برجسته، نمود طراحی در زیستشناسی قابل اعتماد نیست. در نتیجه، هر بینشی که ما را به سمت دیدن طراحی حقیقی در سیستمهای زیستی هدایت کند در حقیقت دارد ما را به سوی بیراههها میبرد.
اما این بینشها الزاما ما را به سوی بیغولهها نمیبرند بلکه میتوانند ما را به راه راست هدایت کنند. در حقیقت، آنها اغلب ما را به حقایقی میرسانند که در صورت نبود این بینشها ما هرگز به این حقایق دست پیدا نمیکردیم. اما دانشمندان چهگونه بین ادراکاتی که ما را به سوی ویرانهها میبرند و آنهایی که ما را به راه راست هدایت میکنند، تفاوت قایل میشوند؟ مشکل علم با شهود این است که غیررسمی و نادقیق است. از این رو دانشمندان برای این که تعیین کنند یک دانستهی شهودی ما را به راه صحیح هدایت میکند یا به سوی ویرانهها، سعی میکنند با تجزیه و تحلیل دقیق و رسمی به آنها واقعیت بخشند. داروینیستها ادعا میکنند که دقیقا همین کار را کردهاند. بنابراین، آنها ادعا میکنند نشان دادهاند که چه موقع ادراکات شهودی ما در مورد طراحی در زیستشناسی شکست میخورد و چهگونه مکانیسم انتخاب داروینی میتواند ساختارهای زیستی در ظاهر طراحی شده را بوجود بیاورد. اما داروینیستها چنین چیزی را نشان ندادهاند. همان طور که در بخشهای قبلی دیدیم، نظریهی داروین هیچ بینشی در مورد ظهور ماشینهای مولکولی پیچیدهی کاهشناپذیر ارایه نمیدهد. در نتیجه ما نیاز داریم که بار دیگر همان شهودهای خودمان را در مورد این که این سیستمها (به خصوص تاژک باکتری) طراحی شدهاند جدی بگیریم. چالش پیش روی نظریهپردازان حامی طراحی این است که آنالیزی رسمی و دقیق فراهم کنند که نشان دهد اداراکات شهودی ما در مورد طراحی در زیستشناسی در واقع توجیهپذیرند و به ویژه نشان دهند که چهطور سیستمهای زیستی مختلف با معیارهایی که من در نوشتههای دیگرم برای تشخیص طراحی توصیف کردهام (معیارهای پیچیدگی مشخص) همخوانی دارند.
اما این تجزیه و تحلیلهای رسمی طراحی محور از سیستمهای پیچیدهی کاهشناپذیر دقیقا چه شکلیاند؟ این تحلیلها چهگونه نشان میدهند که چنین سیستمهایی در واقع هم پیچیده و هم مشخصاند و در نتیجه از معیارهای پیچیدگی مشخص پیروی میکنند و بنابراین طراحی شدهاند؟ جزییات بسیار تخصصی و فنیاند اما منطق کلی که نظریهپردازان طراحی برای نشان دادن پیچیدگی مشخص سیستمهای پیچیدهی کاهشناپذیر به آن استناد میکنند بسیار سرراست است: برای سیستم پیچیدهی کاهشناپذیر مورد نظر و هر پیشساز تکاملی بالقوه، نشان دهید که احتمال این که این پیشساز با مکانیسم داروینی به پیچیدگیکاهشناپذیر رسیده باشد بسیار کوچک است. در چنین تحلیلی، مشخصشدگی هرگز مشکلی خاصی نیست. در چنین مواردی سیستمهای پیچیدهی کاهشناپذیر، هر پیشساز تکاملی و هر چیزی بین پیشساز و سیستم پیچیدهی کاهشناپذیر نهایی همگی همواره برای عملکرد زیستی مخصوص خودشان مشخص شدهاند. همچنین، احتمالات در اینجا باید به طور دقیق محاسبه شوند. تنها کافی است که یک حد بالا برای این احتمالات تعیین کنید و نشان دهید که احتمال محاسبه شده به نسبت این حد کوچک است. بهعلاوه، اگر احتمال تکامل یک پیشساز به یک حدواسط قابل قبول کم باشد، آن وقت احتمال اینکه یک پیشساز بهواسطهی آن حدواسط به یک سیستم پیچیدهی کاهشناپذیر تکامل یافته باشد، به مراتب کوچکتر خواهد بود.
داروینیستها به این رویکرد برای تعیین پیچیدگی مشخص سیستمهای زیستشیمایی پیچیدهی کاهشناپذیر اعتراض دارند. آنها ادعا میکنند تئوریسینهای حامی طراحی، استدلال داروینی را مثل یک استدلال پوشالی از نوع «گردباد در قبرستان ماشینها» ارایه کردهاند. ایدهی «گردباد در قبرستان ماشینها» را اولین بار فرد هویل[38] اخترشناس مطرح کرد. هویل فرض کرد که در یک قبرستان ماشین تمام قطعات مربوط به یک بویینگ 747 در جاهای مختلف پخش و پلا شده است و یک گردباد میآید و همهی قطعات را به نحوی در کنار هم قرار میدهد که در انتها یک بوئینگ 747 آمادهی پرواز باقی میماند]2۸[. داروینیستها اعتراض میکنند که چنین تصویری هیچ ربطی به آن نحوه از تکامل داروینی که این پیچیدگیها را میسازد، ندارد. بر همین اساس، آنها در ساخت سیستمهای پیچیدهی کاهشناپذیری مانند تاژک باکتری هم همان استدلالها را مطرح میکنند تا نشان دهند چنین سیستمهایی نمیتوانند صرفا به شکل تصادفی پدید آمده باشند. اما داروینیستها ادعا میکنند که تکامل در مورد تصادفات نیست بلکه در مورد انتخابطبیعی است که تاثیرات تصادفات را غربال میکند.
مطمئنا اگر حامیان طراحی تنها استدلال میکردند که تصادف محض نمیتواند سیستم پیچیدهی کاهشناپذیر بسازد، آن وقت سزاوار اعتراض «گردباد در قبرستان ماشینها»ی داروینیستها بودند اما این چیزی نیست که نظریهپردازان حامی طراحی میگویند. مشکل تصویر گردباد در قبرستان ماشینهای هویل این است که از دید نظریهی احتمالات، گفتهی هویل سر هم کردن یک بویینگ 747 از اجزای سازندهاش در سختترین حالت ممکن است. اما اگر بخشها همین طور پخش و پلا نبودند چه؟ مثلا، اگر دقیقا همانطور چیده شده بودند که برای ساخت یک بویینگ 747 دارای عملکرد لازم است، چه؟ آیا این به دانش بیشتری نسبت به آنچه که از نظر منطقی به برنامهی شبیهساز تکامل داروینی نسبت داده میشود، نیاز دارد؟
نظریهپردازان طراحی فارغ از تلاش برای دشوار ساختن تکامل سیستمهای پیچیدهی کاهشناپذیری مانند تاژک باکتری، میکوشند تا هرگونه مزیت قانونیای که مکانیسم انتخاب داروینی برای تکامل دادن چنین سیستمهایی نیاز دارد به آن عرضه کنند. یک مزیت که از نظر قانونی نمیتوانیم به مکانیسم انتخاب داروینی بدهیم، دانش پیشین در مورد سیستمی است که تکاملش مورد پرسش قرار گرفته است. این باعث میشود که به مکانیسم داروینی قدرتی غایتشناسانه (در اینجا بینش و برنامه) بدهیم که خود داروین اصرار داشت چنین چیزی وجود ندارد و اگر بنا باشد که نظریهی تکاملی واقعا و به طرزی موثر از طراحی دوری کند، قطعا چنین چیزی امکانپذیر نخواهد بود. با این حال، حتی با سخاوتمندانهترین منافع مشروعی که میتوان به مکانیسم داروینی داد، باز هم احتمال اینکه این مکانیسم بتواند سیستمهای پیچیدهی کاهشناپذیری مانند تاژک باکتری را بسازد، همواره بسیار کم خواهد بود]29[.
دلیل این که این احتمالات همواره بسیار کوچک خواهند شد، دشواربودن هماهنگکردن تغییرات تکاملی متوالی فارغ از غایتشناسی یا هدفمندی است. در مکانیسم داروینی نه انتخاب و نه تغییر بر اساس اهداف آینده انجام نمیشوند (مثل این که در یک باکتری که فاقد تاژک است، هدف این باشد که طی تکامل تاژکی برایش ساخته شود). انتخاب، انتخابطبیعی است که کارش صرفا اعطای منافع آنی به موجود در حال تکامل است. به طور مشابه، دگرگونی و تغییر هم تغییر تصادفی است که کارش این است تا تغییراتی در ساختارهای قابل به ارث رسیدن موجودِ در حال تکامل ایجاد کند، بدون این که در نظر بگیرد چنین تغییراتی برای نسلهای بعد مفید یا مضر است.
مکانیسم داروینی در مسیر تلاش برای هماهنگسازی در ایجاد یک ماشین زیستشیمیایی پیچیدهی کاهشناپذیر که مستلزم تغییرات تکاملی متوالی است، با تعدادی موانع احتمالاتی دلهرهآور روبهرو میشود. از جمله]30[:
1) در دسترس بودن. آیا بخشهایی که مورد نیازند تا یک سیستم زیستشیمیایی پیچیدهی کاهشناپذیر مانند تاژک باکتری را تکامل ببخشند، در دسترساند؟
2) همزمانی. آیا این بخشها در یک زمان در دسترساند تا بتوانند با هم ترکیب شوند و ساختار مورد نظر را بسازند؟
3) تجمع و تمرکز در مکانی خاص: حتی اگر همهی این بخشها در یک زمان برای مشارکت در یک سیستم در حال تکامل در دسترس باشند، آیا این بخشها میتوانند از سیستمی که در حال حاظر در آن ادغام شدهاند جدا شوند و در «جایگاه ساخت» سیستم در حال تکامل حاضر شوند؟
4) تداخل در واکنشهای متقابل. به فرض اینکه قطعات صحیح بتوانند در زمان مناسب در یک مکان گرد هم جمع شوند، چهطور ممکن است بخشهای نادرست را از مداخله در «جایگاه ساخت» سیستم در حال تکامل دور نگاه داشت؟
5) سازگاری تعامل دو جانبه. آیا بخشهایی که به خدمت گرفته شدهاند تا در یک سیستم در حال تکامل گنجانده شوند، سازگاری دو جانبه برای قرار گرفتن در یک شبکه یا ارتباط دو طرفهی تنگاتنگ دارند تا وقتی در کنار هم قرار گرفتند، بتوانند یک سیستم دارای عملکرد یکپارچه را ایجاد کنند؟
6) ترتیب متصل کردن قطعات به هم. حتی اگر همهی قطعات و البته فقط قطعات درست در زمان مناسب به مکان مناسب برسند و حتی اگر سازگاری دو سویه هم وجود داشته باشد، آیا آنها با ترتیب صحیحی کنار هم قرار میگیرند تا یک سیستم عملکردی شکل دهند؟
7) پیکربندی. حتی با قرارگرفتن قطعات صحیح با نظمی صحیح در کنار هم، آیا این قطعات به نحوی صحیح سازماندهی میشوند تا یک سیستم دارای عملکرد بسازد؟
برای این که ببینیم برای از میان برداشتن این موانع چه اقداماتی باید صورت بگیرد، تصور کنید که شما پیمانکاری هستید که برای ساخت یک ساختمان به خدمت گرفته شدهاید. اگر بخواهید با موفقیت این خانه را بسازید باید بر این موانع غلبه کنید. نخست باید معین کنید که تمام مصالح مورد نیاز برای ساخت خانه (برای مثال آجر، الوار چوبی، سیمهای الکتریکی، شیشهها و لولهها) وجود دارند و برای استفاده «در دسترس»اند. سپس باید اطمینان حاصل کنید که میتوانید همهی این مصالح را در یک بازهی زمانی منطقی بدست بیاورید. برای مثال اگر برخی مصالح حیاتی چند سال بعد آماده شوند، شما قادر نخواهید بود خانه را در موعد مقرری که در قرارداد ذکر شده است به پایان برسانید. بنابراین، دسترسی به این مصالح باید به طور مناسبی «همزمان» باشد. سوم، باید همهی این مصالح را به محل ساخت و ساز منتقل کنید. به عبارت دیگر باید تمام مصالح مورد نیاز برای ساخت خانه را به «مکانی» که قرار است خانه در آنجا ساخته شود، منتقل کنید.
چهارم، باید محل ساختوساز را از موادی که ممکن است به خانه آسیب بزنند یا در کار ساخت و ساز اختلال ایجاد کنند پاکسازی کنید. برای مثال وجود مواد زاید رادیواکتیو یا مینهای انفجاری در محل ساخت و ساز به طرز موثری از ساخت یک خانهی قابل استفاده جلوگیری میکند. اگر اندکی از دوز دراماتیک قضیه بکاهیم، فرض کنید مقادیر زیادی از انواع زبالهها و نخالهها (مثلا اشیایی که به ساخت خانه نامرتبطاند مثل قوطی، اسباب بازی شکسته و روزنامه باطله) در محل ساخت و ساز وجود داشته باشند، ممکن است مرتبسازی این شلوغیها و پیدا کردن مصالح ضروری برای ساخت خانه آن قدر سخت شود که خانه هرگز ساخته نشود. این ورود مواد زاید به محل ساخت و ساز و به تعویق افتادن ساخته شدن یک خانهی قابل استفاده را شاید بتوانیم با عنوان « تداخل در واکنشهای متقابل» توصیف کنیم.
پنجم، تهیهی مصالح مورد نیاز برای خانه به طورکلی کافی نیست. به عنوان یک پیمانکار شما همچنین باید اطمینان حاصل کنید که این مصالح به طور مناسبی با همدیگر منطبقاند. بله، نیاز به پیچ و مهره، لوله و زانو و کابل و لوله خرطومیای که کابل را از میان آن رد کنید دارید اما تا وقتی که پیچ و مهره به درستی هماندازه نباشند، تا زمانی که لولهها و زانوها با همدیگر متناسب نباشند و تا زمانی که کابلها درون لولهها جای نگیرند، قادر نخواهید بود که یک خانهی قابل سکونت بسازید. مطمئنا هر بخشی به خودی خود میتواند مصالح ساختمانی بینقصی برای استفاده در این یا آن خانه باشد اما نگرانی شما این یا آن خانه نیست، بلکه خانهای است که خودتان دارید میسازید. تنها زمانی که همهی اجزای موجود در محل ساخت و ساز با همدیگر سازگار باشند و تعاملی صحیح با هم برقرار کنند، شما قادر خواهید بود که خانهای قابل استفاده بسازید. به طور خلاصه، به عنوان یک پیمانکار نیاز است اطمنیان حاصل کنید که نه تنها قطعاتی که به محل ساخت و ساز میبرید همان قطعاتی باشند که به طورکلی مورد نیازند، بلکه باید سازگاری مناسب جهت تعامل دو جانبه هم داشته باشند تا بتوانند به طور موثر کار کنند.
ششم، حتی با وجود همهی مصالح مناسب مورد نیاز در محل ساخت و ساز، باید اطمینان حاصل کنید که این مصالح را به نحوی صحیح در جای خودشان قرار دهید. بنابراین، در ساخت خانه باید ابتدا پیریزی کنید و فونداسیون ساختمان را اجرا کنید. اگر قبل از پیریزی بخواهید دیوارها را بالا ببرید، تلاشتان برای ساخت خانه با شکست مواجه خواهد شد. مصالح درست نیازمند «ترتیب متصل شدن» هم هست تا یک خانهی قابل سکونت ساخته شود. هفتم و آخرین نکته این که مصالح باید به طرزی مناسب سازماندهی شوند. به همین دلیل است که به عنوان پیمانکار بنا، لولهکش و برقکار استخدام میکنید. شما این افراد را صرفا برای متصلکردن مصالح ساختمانی صحیح با نظمی صحیح استخدام نمیکنید بلکه آنها را استخدام میکنید تا این مصالح را به روشی صحیح هم در جایشان قرار دهند. برای مثال، برداشتن آجرها و چیدن آنها با نظم مناسب بسیار خوب است. اما اگر دیوارها با زاویهای عجیب چیده شوند یا اگر دیوار به صورت مورب ساخته شود به طوری که با کوچکترین تکانی سرنگون شود آن وقت حتی اگر همهی ترتیبها هم درست باشد، باز هم نتیجهاش یک خانهی قابل استفاده نخواهد بود. به عبارت دیگر، این که صرفا مصالح مناسب را با ترتیبی صحیح بچینیم کافی نیست، بلکه باید همزمان که مصالح را به هم متصل میکنیم، «پیکربندی» مناسب را هم رعایت کنیم.
اکنون به عنوان یک پیمانکار ساختمان، هیچکدام از این موانع را غیرقابل برطرفکردن نمیبینید، زیرا به عنوان یک عامل هوشمند، میتوانید همهی این وظایف را به طور مناسب هماهنگ کنید تا این موانع را از سر راه بردارید. یک نقشهی معماری برای خانه دارید و میدانید که چه مصالحی برای ساخت خانه مورد نیاز است. میدانید چهطور آنها را تهیه کنید و چهطور در زمان مناسب به مکان مناسب منتقلشان کنید. شما میدانید چهطور می توان این مکان را از شرّ خرابکاران، سارقان، آوار، آب و هوا و هر چیز دیگری که ممکن است تلاشهای شما در ساخت وساز خراب کند، ایمن کنید. میدانید چهطور اطمینان حاصل کنید که مصالح به طور مناسب با یکدیگر منطبق شوند و به همین خاطر وقتی این مصالح کنار هم قرار میگیرند به طرزی صحیح و موثر کار میکنند. ترتیب صحیح چینش آنها را میدانید و با وجود افراد متخصصی که استخدام کردهاید میدانید چهطور این مصالح را در پیکربندیای درست سازماندهی کنید. همهی این «میدانید چهطور»ها از هوش و آگاهی حاصل میشوند و به همین دلیل هم هست که میتوانید یک خانهی قابل استفاده بسازید.
اما مکانیسم تنوع تصادفی داروین و انتخابطبیعی هیچکدام از این «میدانید چهطور»ها را نمیدانند. تنها چیزی که میدانند این است که چهطور به طور تصادفی چیزها را تغییر دهند و سپس آن اصلاحات تصادفیای را که در آن لحظه مفید هستند، حفظ کنند. مکانیسم داروینی یک مکانیسم لذت لحظهای است. اگر مکانیسم داروینی یک پیمانکار ساختمانی بود، ممکن بود برای مزیت آنی دور کردن مزاحمان از محل ساخت و ساز، حتی بدون پیریزی شروع کند به ساختن دیوارها و در نتیجه هیچ وقت خانهی قابل استفادهای نمیتوانست بسازد. این نحوهی کار مکانیسم داروینی است و به همین دلیل هم هست که این قدر محدود است.
تصور کنید برای مکانیسم داروینی برطرفکردن همهی این هفت مانع در راه تکامل تاژک باکتری چه معنایی خواهد داشت. بیاید با یک باکتری شروع میکنیم که نه تاژک دارد، نه ژنی که پروتئینهای تاژک را رمزگذاری کند دارد و نه ژنی که هومولوگ ژنهای کدکنندهی پروتئینهای تاژک باشد. چنین باکتریای باید به مرور زمان به باکتریای تکامل یابد که مجموعهی کاملی از ژنهای مورد نیاز برای گرد هم آوردن یک تاژک کاملا دارای عملکرد را در خود دارد. آیا مکانیسم داروینی کفایت لازم برای هماهنگ کردن همهی رویدادهای زیستشیمیایی مورد نیاز برای برطرف کردن همهی این هفت مانع و در نهایت تکامل تاژک باکتری را دارد؟ پاسخ مثبت به این پرسش نسبت دادن قدرتی خلاقانه به مکانیسم داروینی است که چنین چیزی به طرز فوقالعادهای غیرقابل باور است.
برای بررسی این موضوع بیایید سراغ این هفت مانع برویم، هفت مانعی که هر کدام چالشی بالقوه در راه تکامل داروینی تاژک باکتری است. با «در دسترس بودن» آغاز میکنیم: آیا مکانیسم داروینی میتواند «مانع دسترسی» را برطرف کند؟ برای این کار مکانیسم داروینی باید بتواند یک پروتئین جدید را از همان ابتدا خلق کند (تاژک باکتری، اگر واقعا تکامل یافته باشد، از باکتریای تکامل یافته است که هیچ ژنی، (چه خود ژن و چه هومولوگ آن را) برای ساخت پروتئینهای تشکیل دهندهی تاژک ندارد. درست است که مکانیسم داروینی قادر است پروتئینهای موجود را بازآرایی کند یا آنها را به طور کامل برای وظیفهای جدید به کار گیرد اما هیچ شواهدی در دست نیست که بتواند پروتئینهای مشخص پیچیدهای را از ابتدا خودش بسازد (مشکل پیچیدگیهای مشخص صرفا در سطح ماشینهای مولکولی پیچیدهی کاهشناپذیر بهوجود نمیآید بلکه حتی در سطح پروتئینهایی است که این ماشینها را میسازند و جزء عناصر سازندهی آنها هستند). علاوه بر این، تحقیقات اخیر در مورد حساسیت عملکردی بسیار بالای پروتئینها شواهدی قوی مبنی بر اینکه این طبقه از پروتئینها در اصل به روشهای تدریجی تکاملپذیر نیستند، ارایه میدهند (و در نتیجه همین موانع در راه مکانیسمهای داروینی هم وجود دارد) زیرا انحرافهای کوچک در این پروتئینها همهی عملکردهای زیستی قابل تصور (و نه صرفا عملکردهای زیستی موجود) را نابود میکند]3۱[. بنابراین، بسیار غیرقابل تصور است که مکانیسم داروینی قادر باشد پروتئینهای جدید مورد نیاز برای تکامل تاژک باکتری (و به همین ترتیب ژنهای رمزگذاریکنندهی آنها) را بسازد.
در مورد «مانع همزمانی» چهطور؟ برای مکانیسم داروینی چیرهشدن بر برخی موانع نسبت به سایر موانع آسانتر است و این شاید یکی از آن موانع باشد. انتخابطبیعی میتواند ساختارهایی را که در حال حاضر برخی اهداف زیستی مفید را تامین میکنند نگهدارد. بنابراین، وقتی یک ساختار کارآمد زیستی در دسترس باشد، تمایل بر حفظ و نگهداری آن است. علاوه بر این، برخلاف پیمانکاران ساختمانی که باید پروژه را در بازهی زمانی کوتاه به پایان برسانند، تکامل داروینی هیچ ضربالاجلی برای پایان کارش ندارد (با وجود این دقت داشته باشید که اخترفیزیکدانان یک ضربالاجل طولانی مدت در نظر میگیرند و آن تقریبا پنج میلیارد سال دیگر است؛ زمانی که خورشید تبدیل به یک غول قرمز میشود و این رویداد باعث انبساط خورشید می شود. به این ترتیب خورشید هر چیزی که در راهش قرار میگیرد، از جمله زمین را، میسوزاند]32[. بنابراین به نظر میرسد زمانبندی برای این که ساختارها کی در دسترس سیستمهای در حال تکامل باشد، چندان حیاتی نباشد. تنها چیزی که ممکن است مشکلساز باشد این است که برخی موارد که تا پیش از این عملکرد زیستی مفیدی انجام میدادند و برای تکامل برخی سیستمهای پیچیدهی کاهشناپذیر مورد نیاز هستند، در میانهی فرایندهای تکاملی مزیت عملکردیشان را از دست بدهند و بنابراین بیاستفاده شوند. در چنین مواقعی انتخابطبیعی تمایل دارد این موارد را حذف کند و در نتیجه آن را از دسترس خارج سازد.
از سوی دیگر، به نظر میرسد مکانیسم داروینی برای «مانع تمرکز منابع»[39]، کار بسیار دشواری پیش رو داشته باشد. مشکل در بازبینی مواردی است که قبلا در یک سیستم معین به کار میرفتهاند و حالا باید به شیوهای جدید در سیستم در حال ظهور مورد استفاده قرار گیرند. این سیستمهای در حال ظهور جدید به عنوان سیستمهایی از پیش موجود میکوشند تا با اجزایی که قبلا در سیستمهای دیگر ادغام شده بودند دچار اصلاحات شوند. اما چهقدر محتمل است که بخشی از یک سیستم از آن جدا شود و در محل ساخت یک سیستم از پیش موجود وارد شود و بدین ترتیب آن را به یک سیستم نوظهور با عملکردی جدید یا ارتقا یافته تبدیل کند؟ بهترین شواهد ما پیشنهاد میدهند که این جابهجایی و جایگزینی مواردی که قبلا در سیستمی متفاوت بودهاند عملا نامحتمل است و هر چه تعداد آنها در یک زمان و مکان معین بیشتر شود، از احتمال وقوع آن هم کاسته میشود. دو دلیل برای این موضوع وجود دارد. نخست، محل ساختوساز سیستمهای زیستشیمیایی تمایل دارد که یکپارچگیاش را حفظ کند و تنها به پروتئینهایی اجازهی ورود بدهد که به سیستم مرتبطاند و آنهایی را که ممکن است موجب اختلال شوند، از سیستم دور نگه میدارد. دوم، جدا شدن پروتئینها از سیستمی که در آن مشغولاند امری عادی نیست و برای این کار مجموعهای از تغییرات ژنتیکی مثل مضاعف شدن ژن، تغییرات تنظیمی و جهشهای نقطهای مورد نیاز است.
مانع «تداخل در واکنشهای متقابل»[40]، چالش قبلی در مقابل مکانیسم داروینی را تشدید میکند. اگر تاژک باکتری واقعا حاصل تکامل داروینی باشد آن وقت پیشسازهای تکاملی تاژک باید در این مسیر وجود داشته باشند. این پیشسازها باید به نوبهی خود سیستمهای دارای عملکردی بوده باشند و در مسیرشان برای تکامل به تاژک باید با اجزایی که قبلا کاربردهای دیگری داشتهاند ترکیب شوند و بدین ترتیب متحمل اصلاحات شوند. این اجزا باید در محل ساختوساز همان پیش ساز مورد نظر وجود داشته باشند. اکنون، همانطور که در مورد مانع تمرکز منابع دیدیم، دلیلی وجود ندارد که فکر کنیم چنین چیزی محتمل است. به طور معمول جایگاه ساختوساز یک سیستم زیستشیمیایی معین یکپارچگی خاص خودش را دارد و تنها به پروتئینهای مرتبط با سیستم اجازهی ورود میدهد و پروتئینهای نامرتبطی را که ممکن است مخرب باشند از سیستم دور نگاه میدارد. اما فرض کنید که جایگاه ساختوساز نسبت به پروتئینهای جدید اندکی با رویی گشادهتر برخورد کند (بنابراین مانع مکانی هم تقلیل پیدا کند و احتمال برطرف شدنش افزایش یابد). در این موارد، با استقبال از چیزهایی که میتوانند در تکامل تاژک باکتری کمککننده باشند، جایگاه ساختوساز همچنین از مواردی استقبال میکند که میتوانند مانع تکامل شوند. در نتیجه، هرچه برطرفکردن مانع مکانی آسانتر شود، برداشتن مانع تداخل دشوارتر میشود و بالعکس.
با مانع «سازگاری دو جانبه»[41] ما به سنگینترین چالش پیشروی مکانیسم داروینی میرسیم. مشکل این است که مکانیسم داروینی برای این که بتواند سیستمی را تکامل دهد، باید بخشهایی را که قبلا به منظور خدمت در سایر سیستمها در نظر گرفته شده بودند، بازآرایی کند. اما این همهی ماجرا نیست. برای این کار همچنین لازم است اطمینان حاصل کند که این بخشهای بازآرایی شده به طرز مناسبی با همدیگر ارتباط برقرار میکنند. اگر چنین نشود، سیستم در حال تکامل عملکردش را از دست خواهد داد و بنابراین دیگر مزیتی برای انتخابشدن ارایه نخواهد کرد. محصولات تکامل داروینی هرچه که باشند نتیجهی سرهم شدن قطعات مختلف است. به عبارت دیگر، آنها سیستمهایی هستند که از به هم پیوستن اجزایی که قبلا کاربردهای دیگری داشتهاند، حاصل شدهاند. اگر این اجزا براساس استانداردها و قراردادهایی مشترک ساخته شده بودند، دلیلی وجود داشت تا بخواهیم فکر کنیم که این اجزا میتوانند به طرز موثری با همدیگر کار کنند. اما انتخابطبیعی ظرفیت ایجاد چنین استانداردها و قراردادهایی را ندارد.
خودروهایی را فرض کنید که در چند شرکت اتومبیلسازی مختلف ساخته شدهاند. برای مثال شورولت ایمپالا از ایالات متحده و هوندا آکورد از ژاپن. گرچه این ماشینها تا حدی شبیه هم هستند و زیرسیستمها و بخشهایی دارند که عملکردهای معینی را به طرق معینی انجام میدهند، اما این قسمتها با همدیگر سازگار نیستند. برای مثال، نمیتوانید پیستون یکی از این ماشینها را جدا کنید و به دیگری وصل کنید یا پیچ و مهرهها و بلبرینگهای آنها را با هم جابهجا کنید. این موضوع به این دلیل است که این دو خودرو به طور مستقل و مطابق استانداردها و قراردادهای متفاوتی طراحی شدهاند. البته در کارخانهی شورولت که ایمپالا را ساخته است، استانداردها و قراردادهای مشترکی وجود دارد تا اطمینان حاصل شود که بخشهای مختلف ایمپالا میتوانند به طرزی موثر با هم کار کنند. اما میان دو کارخانهی خودروسازی مختلف (مثلا شورولت و هوندا) هیچ استاندارد و قرارداد مشترکی وجود ندارد (یا خیلی کم وجود دارد) که هنگام ساخت قطعات بخواهند به آنها پایبند باشند. در حقیقت، استانداردها و قراردادهای مشترکی که ارتباط دو جانبهی دو سیستم عملکردی مجزا را تسهیل کند صرفا به طراحی سیستمها بستگی ندارد بلکه به یک طراحی مشترک که به استانداردها و قراردادهای مشترک پاسخگو باشد، بستگی دارد.
اما مکانیسم داروینی فاقد صلاحیت لازم برای چنین طراحی مشترکی است. به عنوان یک مکانیسم ارضای فوری، تنها هدف آن ایجاد ساختارهایی است که مزیتی فوری برای یک موجود در حال تکامل فراهم میکنند و هیچ تعهدی ندارد که اطمینان حاصل کند این ساختارها از قراردادها و استانداردهایی پیروی میکنند که آنها را قادر میسازد تا به طرز موثری با سایر ساختارهای موجود در سیستم کار کنند. بنابراین، فرض کنید مدلی که در بخش چهار برای تکامل تاژک باکتری ارایه شد کاملا دقیق است (هر چند که در این بخش دیدیم این مدل نه دقیق است و نه قابل آزمودن و نه گام به گام). در این صورت، در گامی کلیدی برای تکامل تاژک باکتری، یک پیلوس جابهجا میشود و به سیستم ترشحی نوع III متصل (TTSS) میشود. با وجود این، پیش از این همنشینی، این دو سیستم کاملا مستقل تکامل یافتهاند. متعاقبا، اگر صرفا به یک شانس کور محض استناد کنیم، هیچ دلیلی وجود ندارد که بخواهیم فکر کنیم این سیستمها میتوانند با همدیگر کار کنند؛ درست مانند مثال جابهجا کردن قطعات خودروهایی که به طور مستقل طراحی شده بودند. این ضعف نظریهی داروین را میتوان به صورت تجربی آزمایش کرد: یک TTSS و یک پیلوس دلخواه را در نظر بگیرید و میزان اصلاحات ژنتیکی مورد نیاز را برای این که این پیلوس از میان سیستم توزیع پروتئینی TTSS به بیرون افکنده شود، محاسبه کنید. در حال حاضر، هیچ مدرکی، چه نظری و چه آزمایشگاهی، وجود ندارد که نشان دهد مکانیسم داروینی میتواند مانع سازگاری دو جانبه را برطرف کند.
بر طرف کردن «مانع ترتیب گرد هم آمدن قطعات»[42] هم برای مکانیسم داروینی هدفی بلند پروازانه است. مکانیسم داروینی با رشد پیوسته و اصلاحات متوالی کار میکند؛ این مکانیسم قسمتهای جدید را به سیستمهایی که در حال حاضر دارای عملکرد هستند اضافه میکند و به همین میزان قسمتهایی را که در این سیستمها کار میکنند تغییر میدهد. بدین ترتیب، سیستمهای جدید با عملکردهای جدید یا ارتقا یافتهای شکل میگیرند. حالا توجه کنید که وقتی یک بخش جدید به سیستمی که از پیش دارای عملکرد بوده افزوده میشود، چه اتفاقی میافتد؟ در این مواقع سیستم قبلی تبدیل به یک زیرسیستم ابرسیستم تازه شکل گرفته میشود. به علاوه، ترتیب چینش این زیرسیستم، حداقل در ابتدای کار (قبل انجام اصلاحات بعدی)، همان خواهد بود که وقتی این زیرسیستم، خودش سیستم مستقلی بود. در کل، صرف این که بخشهای یک زیرسیستم میتوانند در یک نظم معین کنار همدیگر قرار بگیرند به این معنا نیست که این بخشها وقتی در یک ابرسیستم گنجانده شدند، باز هم میتوانند با همان نظم کنار هم قرار بگیرند. در حقیقت، در تکامل سیستمهایی مانند تاژک باکتری، میتوانیم انتظار داشته باشیم که نظم چیدمان قسمتهای مختلف دستخوش جایگشتهایی اساسی شود. اما این نظم چهگونه دقیقا به همان جایگشتی که باید در میآید؟ برای بیشتر سیستمها ترتیب چینش تثبیت شده است و سیستم اجازهی انحرافات قابل توجه را نمیدهد. مسئولیت ارایهی شواهد بر دوش داورینیستها است که نشان دهند در یک سیستم در حال تکامل مکانیسم داروینی نه تنها ظهور بخشهای صحیح را هماهنگ میکند بلکه ترتیب چینش آنها را هم تعیین و هماهنگ میکند. داروینیستها هیچ کار این چنینیای نکردهاند.
سرانجام، به «مانع پیکربندی»[43]میپردازیم. در طراحی و ساخت مصنوعات دست انسان، این مانع سختترین مانع برای رفع و رجوع کردن است. با این حال، در تکامل سیستمهای زیستشیمیایی پیچیدهی کاهشناپذیری مانند تاژک باکتری، این یکی از آسانترین موانع برای برطرف کردن است، به این دلیل که در تاژک و سیستمهای این چنینی قطعات زیستشیمیایی به صورت تصادفی و بینظم و ترتیب کنار هم قرار نمیگیرند. در عوض، زمانی که برخوردهای تصادفی اجازه دهد که هر بار یکی از قطعات تاژکی برهمکنشهای خاص، تعاونی و الکترواستاتیک موضعی برای در هم تنیده شدن برقرار کند، آنگاه این قطعات میتوانند با پیکربندی مناسب در جای خود قرار بگیرند. بنابراین، در تکامل تاژک باکتری، به محض این که موانع سازگاری دو جانبه و ترتیب سر هم شدن قطعات بر طرف شود، مانع پیکربندی هم برطرف میشود. یک اصل کلی در اینجا وجود دارد: برای ساختارهای خود سازمان دهندهای مانند سیستمهای زیستی، پیکربندی یک محصول فرعی از سایر محدودیتها است (مانند سازگاری دو جانبه و ترتیب سر هم شدن قطعات). اما توجه داشته باشید این به آن معنا نیست که پیکربندی این سیستمها کاملا مجانی به دست میآید، بلکه به این معنا است که هزینهی پیکربندی آنها در هزینهی سایر موانع گنجانده شده است.
این هفت مانعی را که توضیح دادم نباید صرفا مجموعهای از چالشهای ذهنی یا کیفی محض در نظر گرفت. میتوان به صورت ذهنی و کیفی چالشی را که این منابع در راه مکانیسم داروینی میگذارند، تعیین کرد. همراه با هر مانع یک احتمال وجود دارد:
احتمالدسترسی احتمال این که انواع قسمتهای مورد نیاز برای تکامل یک سیستم زیستشیمیایی پیچیدهی کاهشناپذیر معین در دسترس باشند.
احتمالهمزمانی احتمال این که همهی قسمتها در زمان مناسبی در دسترس باشند که بتوان در صورت نیاز آنها را در سیستم در حال تکامل گنجاند.
احتمالمکانیابی احتمال این که این بخشها، که در زمان مناسب در دسترس هستند بتوانند از سیستمی که در حال حاضر در آن دخیلاند جدا شوند و در مکان مناسب برای گرد آمدن در سیستم جدید تجمع پیدا کنند.
احتمالتداخل احتمال این که سایر قسمتهایی که امکان دارد موجب تداخل در واکنشهای متقابل شوند و در نتیجه مانع از شکلگیری سیستمهای پیچیده کاهشناپذیر مورد نظر شوند، از محلی که سیستم در حال سر هم شدن است دور شوند.
احتمالسازگاری احتمال این که بخشهای به خدمت گرفته شده برای گنجانده شدن در یک سیستم در حال تکامل با همدیگر سازگاری دو جانبه داشته باشند و در نتیجه بتوانند با هم یک سیستم دارای عملکرد را شکل بدهند.
احتمالترتیب احتمال این که حتی قطعات صحیحی که در زمان مناسب در مکانی مناسب قرار دارند و حتی به طور کامل سازگاری دو جانبه دارند، بتوانند با ترتیبی صحیح به هم متصل شوند و تشکیل یک سیستم دارای عملکرد را بدهند.
احتمالپیکربندی احتمال این که حتی اگر همهی قطعات با ترتیبی صحیح گرد هم آمده باشند، سازماندهی فضاییشان به طریقی باشد که بتواند یک سیستم دارای عملکرد بسازد.
توجه داشته باشید که هر کدام از این احتمالات منوط به احتمال قبلیاست. بنابراین، احتمال همزمانی، احتمال همزمان در دسترس بودن قطعات را به شرطی تعیین میکند که همهی قطعات در عمل در دسترس باشند. بنابراین به شرطی میتوان احتمال ترتیب سر هم شدن قطعات را تعیین کرد که همهی قطعات در زمانی مناسب (احتمال همزمانی) در مکان صحیح (احتمال مکانیابی) بدون مزاحمت واکنشهای متقابل و با سازگاری دو جانبهی کامل در دسترس باشند (احتمال دسترسی). به عنوان نتیجه، احتمال شکل گیری یک سیستم پیچیدهی کاهشناپذیر به واسطهی مکانیسمهای داروینی نمیتواند از حاصل زیر بیشتر باشد (توجه داشته باشیداز آنجا که هر کدام از احتمالات منوط به احتمال قبلی است، در به دست آوردن این حاصل امکان از قلم انداختن هیچ کدام از احتمالات مستقل وجود ندارد):
احتمالپیکربندی× احتمالترتیب× احتمالسازگاری× احتمالتداخل× احتمالمکانیابی× احتمالهمزمانی× احتمالدسترسی
اگر ما اینجا احتمالمنشا را به عنوان احتمال این که یک سیستم پیچیدهی کاهشناپذیر از مکانیسمهای داروینی نشات گرفته باشد تعریف کنیم، آنگاه به نامعادلهی زیر میرسیم:
احتمالپیکربندی × احتمالترتیب × احتمالسازگاری × احتمالتداخل × احتمالمکانیابی × احتمالهمزمانی × احتمالدسترسی≥ احتمالمنشا ]33[
نامعادلهی منشا پیامدهای بسیار دور از دسترسی دارد. از آنجا که احتمالها اعدادی بین صفر و یک هستند، این نامعادله به ما میگوید حتی اگر فقط یکی از احتمالات سمت راست کوچک باشد، آن وقت احتمال منشا هم کوچک خواهد بود (در واقع، بزرگتر از هیچ کدام از احتمالات سمت راست نخواهد بود). نتیجه این میشود که لازم نیست همهی احتمالات سمت راست را حساب کنیم تا اطمینان حاصل کنیم که احتمال منشا کوچک است. همچنین، نتیجهی دیگر این است که هیچکدام از این احتمالات نیازی به محاسبهی دقیق ندارند. اگر حد هر کدام از موارد بالا کوچک باشد، آنگاه احتمال مربوط به آن هم کوچک است در نتیجه احتمال منشا هم کوچک است و اگر احتمال منشا کوچک باشد آن وقت سیستم پیچیدهی کاهشناپذیر مورد نظر هم نامحتمل خواهد بود و هم مشخص (همهی این سیستمهای پیچیدهی کاهشناپذیر برای عملکرد زیستیای خودشان مشخص شدهاند). این نکته به این نتیجه منتهی میشود که اگر احتمال منشا کوچک باشد، آن وقت سیستم مورد نظر پیچیدگی مشخص از خود نشان میدهد و از آنجا که پیچیدگی مشخص یک مشخصهی تجربی قابل توجه برای طراحی است، در نتیجه خود سیستم هم طراحی شده است.
تصور میکنم مقایسهی احتمال منشا با معادلهی دریک[44] کمک کننده است؛ معادلهای که در جریان جستوجو برای هوش فرازمینی به دست آمد. در سال 1960، اخترفیزیکدانی به نام فرانک دریک اولین کنفرانس جستوجو برای هوش فرازمینی را ترتیب داد و معادلهای را که حالا به معادلهی دریک مشهور است، در آنجا معرفی کرد.
[34] N = N* x fp x ne x fl x fi x fc x fL
حروف معادله برای مفاهیم زیر بکار میروند:
= N تعداد تمدنهای فرازمینی که میتوانیم با آنها تماس برقرار کنیم
= N*نرخ متوسط تشکیل ستارهها در کهکشان راهشیری
= fp نسبتی از این ستارهها که دارای سیاره هستند
= ne تعداد متوسط سیاراتی که احتمال شکلگیری حیات در آنها وجود دارد
= fℓ نسبت سیارههایی که در طول عمرشان عملا حیات در آن شکل میگیرد به تعداد سیاراتی که احتمال شکلگیری حیات در آنها وجود دارد
= fi نسبت سیارههای دارای حیات که میتوانند حیات هوشمند (تمدن) داشته باشند
= fc نسبت تمدنهای دارای فناوری با علایمی قابل دریافت در کهکشان
fL = مدت زمانی که آن تمدنها قابلیت برقراری تماس داشته باشند (طول عمر تمدن)
معادلهی دریک مقیاسی است برای تعیین احتمال موفقیت جستوجوها برای یافتن هوش فرازمینی: N بزرگتر، نشان میدهد احتمال بیشتری وجود دارد که محققان بتوانند در فضای دوردست نشانهای از هوش و آگاهی پیدا کنند.
درست مانند نامعادلهی منشا، هفت عبارت معادلهی دریک را تعیین میکنند، هفت عبارتی که سمت راست معادله قرار دارند. علاوه بر این، هر کدام از این هفت عبارت مانند عبارات نامعادلهی منشا به طور متوالی به همدیگر وابستهاند. برای مثال، نسبت سیاراتی که میتوانند حیات هوشمند تکامل دهند در پرتو نسبت سیاراتی که میتوانند حیات سادهتر را تکامل دهند قابل تعریف است.
به رغم شباهتهایی که بین معادلهی دریک و نامعادلهی منشا – به خصوص این که هر دو به دنبال نشانهای از هوش و آگاهی میگردند – وجود دارد، این دو یک تفاوت آشکار با هم دارند. برای این که معادلهی دریک بتواند ما را متقاعد کند که جستوجو برای هوش فرازمینی احتمالا موفقیتآمیز است، «هیچ کدام از عبارات» سمت راست معادله نباید بسیار کوچک باشند. تنها در این صورت است که محققان شانسی منطقی برای کشف نشانههای هوش فرازمینی خواهند داشت. اما در نامعادلهی منشا، برای تضمین پیچیدگی مشخص و در نتیجه طراحی یک سیستم پیچیدهی کاهشناپذیر، تنها کافی است نشان دهیم که «یکی از عبارات» سمت راست نامعادله به اندازهی کافی کوچک است. با توجه به کاربردهای عملی این فرمولها، این تفاوت همهی تفاوتها را در جهان رقم میزند.
مشکل معادلهی دریک این است که بیشتر عبارات آن را نمیتوان تخمین زد. همانطور که مایکل کریشتون[45] در یک سخنرانی عمومی در موسسهی تکنولوژی کالیفرنیا که به شکل وسیعی منتشر شد، میگوید:
تنها راهی که این معادله بتواند کار کند این است که جای عبارات مختلف را با حدسیات پر کنیم و همانطور که برای همگان روشن است، حدسیات صرفا بیان تعصبات ما هستند. هیچ «حدس آگاهانهای» وجود ندارد. هیچ راهی نیست که طی آن بتوانیم با حدسی آگاهانه تعیین کنیم چند سیاره دارای حیات با توانایی برقراری تماس وجود دارد. صرفا پیش داوری و تعصب است. به عنوان نتیجه، معادلهی دریک میتواند مقادیری بین «میلیاردها میلیارد» و صفر داشته باشد. عبارتی که بتوان هر معنایی را از آن برداشت کرد، هیچ معنایی ندارد. من عمیقا باور دارم که علم تنها به فرضیههای قابل آزمودن تن میدهد. معادلهی دریک قابل آزمودن نیست. حتی ذرهای شواهد وجود ندارد که نشان دهد نوع دیگری از حیات وجود دارد و طی چهل سال تحقیقات هم چیزی حاصل نشده است]35[.
منظور کریشتون از قابل آزمودن این است که بتوان عبارات معادلهی دریک را با اطمینان کافی تخمین زد تا بتوان وجود هوش فرازمینی را تایید کرد. مطابق مطالب گفته شده، زمانی معادلهی دریک میتواند حضور هوش فرازمینی را تایید که بتوان به طرزی مستدل همهی عبارات آن را تخمین زد (که در حال حاضر چنین چیزی ممکن نیست) و همچنین معلوم شود که این عبارات مقادیر بزرگی دارند]36[. در مقابل، اگر در نامعادلهی منشا تنها یکی از عبارات به طرزی مستدل تخمین زده شود و معلوم شود که مقادیر کوچکی دارد آنگاه این نامعادله میتواند تایید کند که یک آگاهی در شکلگیری ساختارهای زیستی پیچیدهی کاهشناپذیر دخیل است. این به این خاطر است که به محض این که معلوم شود یکی از عبارات سمت راست نامعادلهی منشا کوچک است، احتمال منشا هم الزاما کوچک میشود.
هیچ یک از عبارات نامعادلهی منشا به صورت تئوری قابل آزمودن نیست. برای مثال احتمال سازگاری دو جانبه را در نظر بگیرید. سیستمهای زیستشیمیایی موجود میتوانند به هم بپیوندند (هر سیستمی از پروتئینهای منفرد گرفته تا ماشینهای زیستشیمیایی پیچیده) و با آزمایشهای تجربی میزان سازگاری دو جانبه آنها را اندازه گرفت. همچنین میتوان سیستمهای زیستشیمیاییای را که در حال حاضر وجود دارند برداشت و انحرافی در آنها به وجود آورد و سپس آنها را سر جایشان برگرداند. به میزانی که این سیستمها میتوانند انحراف را تحمل کنند، با روشهای داروینی هم قابل تکاملاند. آزمایشهایی مانند این را میتوان روی سیستمهای زیستشیمیایی واقعی هم پیاده کرد. حتی میتوان روی فرایندهای زیستشیمیایی شبیهسازی شده در کامپیوتر هم اجرا کرد. نکته این است که با احتمال سازگاری دو جانبه و سایر احتمالات موجود در نامعادلهی منشا، هیچ مانع ذاتیای برای برآورد یک تخمین قابل اعتماد و تایید شده وجود ندارد. هم داروینیستها و هم نظریهپردازان طراحی تمایل زیادی برای محاسبهی این احتمالات دارند. تحقیقاتی در این زمینه آغاز شده که تازه در آغاز راه قرار دارند.
نامعادلهی منشا هیچ تعصب ذاتیای ندارد و تلاش نمیکند با پیشداوری حکم کند که یک سیستم زیستشیمیایی پیچیدهی کاهشناپذیر طراحی شده است. تا زمانی که احتمالات بزرگ باشند یا محاسبه نشده باشند، پیش فرض بر عدم پیچیدهی مشخص بودن سیستم و در نتیجه علیه طراحی است. از سوی دیگر، اگر معلوم شود که تنها یکی از این احتمالات مقداری دارد که به طرز چشمگیری کوچک است، آنگاه این پیش فرض به سمت پیچیدگی مشخص میچرخد و فرض بر این میشود که سیستم طراحی شده است. در این حالت، نامعادلهی منشا شرایط بازی برابری برای انتخاب بین طراحیهوشمندانه و نظریهی داروین فراهم میکند. داروینیستها هر بار که برای پشتیبانی از نظریهشان به رویدادهای با احتمال زیاد رجوع میکنند، در حقیقت به طور ناخودآگاه و ضمنی موافقت خودشان را با نامعادلهی منشا اعلام میدارند. برای مثال، در جستوجو برای تایید اینکه مقاومت باکتریها به آنتیبیوتیک در اثر مکانیسم داروینی حاصل میشود و نه طراحی، داروینیستها با خوشحالی اشاره میکنند که احتمال جهشهای نقطهای مورد نیاز برای مقاومت به آنتیبیوتیک بسیار بالا است.
اما با وجود این که با هر بار تایید نظریهی داروین این نامعادله هم به طور ضمنی تایید میشود، داروینیستها به سرعت انکار میکنند که این نامعادله بتواند قانونا برای رد نظریهی داروین مورد استفاده قرار گیرد. این استاندارد دوگانه دقیقا رویکرد خود داروین است. او در منشا انواع چالش زیر را مطرح کرد که «اگر هر گونه اندام پیچیدهای وجود داشته باشد که طی اصلاحات متعدد، متوالی و جزیی بهوجود نیامده باشد، آنگاه نظریهی من به طور قطع شکست میخورد. اما من نتوانستهام چنین موردی بیابم.»]37[.
کاری که داروین در اینجا دارد انجام میدهد مثل این است که بگوید اگر شیر آمد من میبرم و اگر خط آمد تو میبازی. در واقع، چالش او اصلا چالش نیست. تنها تضمین میکند که نظریهاش هرگز مورد تحقیقات منتقدان قرار نخواهد گرفت و نمیتواند قرار بگیرد. همان طور که رابرت کونز[46] اشاره میکند:
چهطور میتوان ثابت کرد که چیزی با روندی غیر از روند «اصلاحات متعدد، متوالی و جزیی» شکل گرفته است؟ و اصلا چرا منتقدان [نظریهی داروین] باید چنین کاری بکنند؟ مسئولیت این کار بر دوش داروین و مدافعانش است که حداقل چند اندام پیچیده نشان دهند که در طبیعت با همین روندی که میگویند یعنی مجموعهای مفصل از اصلاحات جزیی، شکل گرفته باشند ]3۸[.
حتی برای استفاده از نامعادلهی منشا هم نخست باید مسیرهای تکاملی خاصی در دست داشت که به سیستمهای زیستشیمیایی پیچیدهی کاهشناپذیری مثل تاژک باکتری ختم میشوند. تنها با داشتن چنین پیشنهادی است که میتوان احتمالاتی را که در نامعادله ظاهر میشوند تخمین زد. بهعلاوه، همواره یک روند ثابت در مورد این نمونهها وجود دارد؛ این که این نمونهها همیشه نشان از بیکفایتی مکانیسم داروینی دارند چون تا امروز ثابت شده که احتمالات نامعادلهی منشا سیستمهای زیستشیمیایی پیچیدهی کاهشناپذیر همواره کوچک است. نظریهپردازان طراحی کوچک بودن این احتمالات را به عنوان تاییدی قوی بر مشخص بودن پیچیدگی این سیستمها و در نتیجه نشانهای بر طراحی بودنشان میدانند. در مقابل، داروینیستها میگویند که احتمالات پایین صرفا نشان میدهد که زیستشناسی تکاملی هنوز راه زیادی تا یافتن مسیرهای تکاملیای دارد که طی آنها این سیستمهای مورد نظر ساخته میشوند.
حق با کیست؟ آن چه فعلا مشهود است این است که هیچکدام از طرفین حاضر نیستند به این زودی از موضع خود کوتاه بیایند. از دید نظریهپردازان طراحی، داروینیستها به طرزی مصنوعی نظریهی داروین را ایزوله کردهاند و آن را در مقابل رد شدن اصولش ایمن کردهاند چون جهان ناشناختههای مسیر داروینی هرگز از پای در نمیآید. از سوی دیگر، تنهای چیزی که داروینیستها را راضی میکند تا بار اثبات را به سوی داروینیسم بگردانند این است که تمام مسیرهای قابل تصور داروین از اساس رد شوند. از دید یک ناظر بیرونی که تعصبی در مورد نتیجهی بحث ندارد، عدم توازن این موضعگیریها بدیهی خواهد بود. طراحیهوشمندانه این امکان را فراهم میکند تا شواهد زیستی بتوانند آن را رد یا تایید کنند. داورینیسم اما در مقابل، هیچ بار اثباتی را بر عهده نمیگیرد و خودش را به طور پیش فرض به عنوان برنده در مقابل طراحیهوشمندانه در نظر میگیرد.
این عدم تمایل داروینیسم در پذیرفتن بار اثباتش شایستهی علم نیست. علم، اگر قرار است تحقیقاتی نامتعصبانه در طبیعت باشد، باید به تمام طیفهای توضیحات منطقی محتمل به یک اندازه اجازهی موفقیت بدهد. به ویژه علم نباید با احکام دلبخواه احتمالات منطقی را از دور خارج کند. زیستشناسی تکاملی، با دادن امتیازات نامنصفانه به استدلال داروینی، انگار از قبل این مسئله را حل کرده است و بدون در نظر گرفتن شواهد تجربی حکم صادر کرده است که کدام توضیح درست است و کدام غلط. این علم نیست، یک فلسفهی بیپشتوانه است. حتی اگر طراحیهوشمندانه نظریهی درستی در مورد منشا حیات نباشد، تنها راهی که علم میتواند این موضوع را کشف کند این است که طراحی را به عنوان یکی از احتمالات زنده بپذیرد نه این که آن را از پیش حذف کند. داروین به طرزی ناعادلانه صحنه را به سود نظریهی خودش چیده است. او در منشا انواع مینویسد: «یک نتیجهی منصفانه تنها با بررسی و سبک سنگین کردن حقایق و استدلالهای دو سوی هر پرسش بهدست میآید»]3۹[. اکنون اما کفهی ترازو از سمت داروینیسم به سوی طراحی سنگینی میکند.
یادداشتها:
- Bruce Alberts, “The Cell as a Collection of Protein Machines: Preparing the Next Generation of Molecular Biologists,” Cell 92 (8,February 1998): 291
- Adam Wilkins, “A Special Issue on Molecular Machines,” BioEssays 25(12) (December 2003): 1146.
- ویلیام دمبسکی در این تعریف از پیچیدگی کاهش ناپذیر، تعریف مایکل بیهی را اصلاح و تعمیم داده است. برای مشاهدهی تعریف اصلی بیهی نگاه کنید به: Darwin’s Black Box: The Biochemical Challenge to Evolution (New York: Free Press, 1996), 39 برای مشاهدهی اصلاحات و تعمیم دمبسکی نگاه کنید به: No Free Lunch: Why Specified Complexity Cannot Be Purchased without Intelligence (Lanham, Md.: Rowman and Littlefield, 2002), sec. 5.9.
- Charles Darwin, On the Origin of Species, facsimile 1st ed. (1859; reprintedCambridge, Mass.: Harvard University Press, 1964), 189
- نگاه کنید به:Howard C. Berg, Random Walks in Biology, exp. ed. (Princeton: Princeton University Press, 1993), 134
برگ در این باره مینویسد: «اشریشیا کولی گیرندههایی برای اکسیژن و همچنین گیرندههایی برای الکترون، شکرها، آمینواسیدها و دیپپتیدها دارد و اشغال بودن این گیرندهها را به عنوان تابعی از زمان تحت نظارت قرار میدهد. احتمال این که سلول با سرعت زیاد و در مسیری مستقیم حرکت کند (چرخیدن تاژک خلاف جهت عقربهی ساعت) یا به صورت تصادفی و آشفته حرکت کند (چرخیدن تاژکهایش در جهت عقربههای ساعت) به مدت زمان اشغال این گیرندهها بستگی دارد. ما بر اساس پاسخهایی که سلولها به پالسهای کوچک مواد شیمیاییای که با میکروپیپتها به آنها داده میشود، میدهند، میدانیم که این اندازهگیریها حدود 4 ثانیه طول میکشد. یک سلول مدت زمانی را که یک گیرندهی خاص– تنها گیرندهای که این مطالعات روی آن انجام شده، گیرندهی آسپارتات است- در ثانیهی قبل اشغال کرده با مدت زمان اشغال بودن در سه ثانیهی قبل مقایسه میکند و به این تفاوت پاسخ میدهد. حال با توجه به توزیع دورانی، دورهی خروج اشریشیا کولی 60 درجه در چهار ثانیه است. اگر اندازهگیری تفاوت غلظتها بیش از این طول بکشد، سلول آن را اعمال نخواهد کرد چون پیش از اعمال نتایج دوره را تغییر خواهد داد. از سوی دیگر، اگر این اندازهگیریها در یک مقیاس زمانی بیش از حد کوچک به پایان برسد، دقت آنها کافی نخواهد بود. ای. کولی تعداد مولکولها را بر اساس انتشار آنها به گیرندههایشان میشمرد و این کاری زمان بر است. خطای نسبی (انحراف معیار تقسیم بر میانگین) با مجذور مقدار شمارش شده کاهش مییابد. بنابراین، در تصمیم برای بهتر یا بدتر شدن حیات ای. کولی با توجه حرکت چرخشی براونی از حداکثر زمان ممکن بهره میبرد.
- Michael Behe, Darwin’s Black Box (New York: Free Press, 1996), 69–73.
- John Postgate, The Outer Reaches of Life (Cambridge: Cambridge University Press, 1994), 160.
- For reviews in the popular press see James Shreeve, “Design for Living,” New YorkTimes, Book Review Section (4 August 1996): 8; Paul R. Gross, “The Dissent of Man,”Wall Street Journal (30 July 1996): A12; and Boyce Rensberger, “How ScienceResponds When Creationists Criticize Evolution,” Washington Post (8 January 1997):
H01. For reviews in the scientific journals see Jerry A. Coyne, “God in the Details,” Nature 383 (19 September 1996): 227–228; Neil W. Blackstone, “Argumentum Ad Ignorantiam,” Quarterly Review of Biology 72(4) (December 1997): 445–447; and Thomas Cavalier-Smith, “The Blind Biochemist,” Trends in Ecology and Evolution 12 (1997): 162–163 - .نگاه کنید به: صفحهی جان کاتالنو با عنوان «جعبه سیاه داروین» در http://www.world-ofdawkins.com/Catalano/box/behe.htm (last accessed October 21, 2003)
- Franklin Harold, The Way of the Cell: Molecules, Organisms and the Order of Life (Oxford: Oxford University Press, 2001), 205. James Shapiro, “In the Details … What?” National Review (16 September 1996): 62–65′
- .Darwin, Origin of Species, 82
- Thomas D. Schneider, “Evolution of Biological Information,” Nucleic Acids Research 28(14) (2000): 2794
- همان
- Darwin, Origin of Species, 189
- H. Allen Orr, “Darwin v. Intelligent Design (Again),” Boston Review (December/January 1996-1997): 29
- همان
- نگاه کنید به:Nguyen, I. T. Paulsen, J. Tchieu, C. J. Hueck, M. H. Saier Jr., “Phylogenetic Analyses of the Constituents of Type III Protein Secretion Systems,” Journal of Molecular Microbiology and Biotechnology 2(2) (2000):125–44
- Kenneth R. Miller, “The Flagellum Unspun: The Collapse of ‘Irreducible Complexity’,” in W. Dembski and M. Ruse, eds., Debating Design: From Darwin to DNA (Cambridge: Cambridge University Press, forthcoming 2004)
- Ian Musgrave, “Evolution of the Bacterial Flagellum,” in M. Young and T. Edis, eds., Why Intelligent Design Fails: A Scientific Critique of the New Creationism (Piscataway, N.J.: Rutgers University Press, forthcoming 2004)
- Nicholas Matzke, “Evolution in (Brownian) Space: A Model for the Origin of the Bacterial Flagellum,” published online at http://www.talkreason.org/articles/flag.pdf (last accessed December 1, 2003)
- همان
- See Kenneth R. Miller, Finding Darwin’s God (New York: HarperCollins, 1999), ch. 5.
- Harold, Way of the Cell, 205.
- Lynn Margulis and Dorion Sagan, Acquiring Genomes: A Theory of the Origins of Species (New York: Basic Books, 2002), 103
- William Dembski, The Design Inference: Eliminating Chance through Small Probabilities (Cambridge: Cambridge University Press, 1998); William Dembski, No Free Lunch: Why Specified Complexity Cannot Be Purchased without Intelligence (Lanham, Md.: Rowman and Littlefield, 2002)
- Richard Dawkins, The Blind Watchmaker (New York: Norton, 1987), 1
- Francis Crick, What Mad Pursuit (New York: Basic Books, 1988), 138.
- Fred Hoyle, The Intelligent Universe (New York: Holt, Reinhart, and Winston, 1984), 19
- See, for instance, Dembski, No Free Lunch, sec. 5.10.
- همان، همچنین نگاه کنید به:
Angus Menuge, Agents Under Fire: Materialism and the Rationality of Science (Lanham, Md.: Rowman and Littlefield, forthcoming 2004), ch. 4.
- Douglas Axe, “Extreme Functional Sensitivity to Conservative Amino Acid Changes on Enzyme Exteriors,” Journal of Molecular Biology 301 (2000): 585–595
- Hubert P. Yockey, Information Theory and Molecular Biology (Cambridge: Cambridge University Press, 1992), 220–221.
- توجه داشته باشید که نیازی نیست این نامعادله به شکل یک معادلهی قطعی در آید زیرا هر زمان که بخواهیم میتوانیم آن را با عبارات اضافی اصلاح کنیم. برای مثال، یک احتمال اضافی فرض کنید به اسم «احتمال نگهداری»؛ احتمال این که قطعاتی که در زمان مناسب در مکان مناسب در دسترساند تا زمانی که تاژک باکتری به طور کامل ساخته شود در همان جا باقی بمانند. احتمال نگهداری بدین ترتیب به احتمال دسترسی، همزمانی و موقعیتی وابسته است و میتواند به عنوان ضریبی بعد از این عبارات در نامعادله گنجانده شود.
- Carl Sagan, Cosmos (New York: Random House, 1980), 299
- Michael Crichton, “Aliens Cause Global Warming,” Caltech Michelin Lecture, January 17, 2003, available online at http://www.crichton-official.com/speeches/speeches_ quote04.html (last accessed January 3, 2004)
- توجه داشته باشید که معادلهی دریک در حال حاضر میتواند وجود هوش فرازمینی را رد کند: تنها کافی است صرفا یکی از عبارات آن را به طرزی مستدل تخمین زد و نشان داد که این عبارت مقدار کوچکی دارد. نتیجه این که با معادلهی دریک آزمودن احتمال شکست هوش فرازمینی بسیار آسانتر از احتمال موفقیت آن است.
- Darwin, Origin of Species, 189.
- Robert Koons, “The Check Is in the Mail: Why Darwinism Fails to Inspire Confidence,” in W. A. Dembski, ed., Uncommon Dissent: Intellectuals Who Find Darwinism Unconvincing (Wilmington, Del.: ISI Books, 2004)
- Darwin, Origin of Species, 2.
منبع:
http://www.designinference.com/documents/2004.01.Irred_Compl_Revisited.pdf
[1] Irreducible Complexity
[2] Darwin’s Black Box
[3] Cell
[4] Bruce Alberts
[5] National Academy of Sciences
[6] The Cell as a Collection of Protein Machine
[7] Bio Essays
[8] Adam Wilkins
[9] Lehigh University
[10] Darwin’s Black Box
[11] Irreducible complexity
[12] irreducible core
[13] cumulative complexity
[14] Michael Behe
[15] Darwin’s Black Box
[16] Howard Berg
[17] John Postgate
[18] argument from irreducible complexity
[19] ergodic theory
[20] coevolution
[21] leprechauns
[22] Franklin Harold
[23] James Shapiro
[24] causal adequacy
[25] Thomas Schneider
[26] patchworks
[27] Allen Orr
[28] University of Rochester
[29] Yersinia pestis
[30] pilus
[31] Franklin Harold
[32] Lynn Margulis
[33] The Design Inference
[34] No Free Lunch
[35] specified complexity
[36] Richard Dawkins
[37] Francis Crick
[38] Fred Hoyle
[39] localization hurdle
[40] interfering cross-reaction
[41] interface-compatibility hurdle
[42] order-of-assembly hurdle
[43] configuration hurdle
[44] Drake equation
[45] Michael Crichton
[46] Robert Koons



