
ماشینهای مولکولی: پشتوانهی تجربی برای استنباط طراحیمایکل بیهی/ ترجمه: امیرحسین لطیفی

مجموعهی چشمها
چهگونه میبینیم؟ در قرن نوزده آناتومی چشم با جزییات کامل شناسایی شد. مکانیسم پیچیدهای که چشم به کار میگیرد تا تصویری دقیق از دنیای بیرون به ما ارایه دهد، همهی کسانی را که در این تحقیقات دست داشتند، مبهوت کرد. دانشمندان قرن نوزدهم به درستی نشان دادند که اگر کسی آنقدر بدشانس باشد که یکی از بسیار ویژگیهای یکپارچهی چشم، مانند عدسی، عنبیه یا عضلات ماهیچهای را از دست بدهد، نتیجهی اجتنابناپذیر آن کاهش شدید بینایی یا نابینایی کامل خواهد بود. از همین رو آنها نتیجه گرفتند که چشم تنها در صورتی که کاملا سالم و تمام و کمال باشد میتواند کارکرد صحیح داشته باشد.
چارلز داروین در راستای بررسی ایرادات احتمالیای که ممکن بود به نظریهی فرگشت براساس انتخاب طبیعی وارد شود، در کتاب منشا انواع[1] خود در فصلی با عنوان «اندامهای پیچیده و در اوج کامل» به بحث در مورد مشکل چشم میپردازد. او پی برد که ظهور ناگهانی اندامی به پیچیدگی چشم در یک نسل رویدادی معادل یک معجزه است. برای اینکه فرگشت داروینی باورپذیر باشد، باید مشکلی که عموم مردم در تجسم شکلگیری اندامهای پیچیده داشتند به نوعی برطرف میشد.
داروین به زیرکی از پس این کار برآمد. او برای این کار به تبیین مسیری که احتمالا فرگشت استفاده کرده است تا چشم را بسازد نپرداخت، بلکه در عوض به انواعی از جانوران اشاره کرد که ساختارهای چشمی متنوع داشتند: از سادهترین لکههای گیرندهی نور تا چشمهای دوربینی پیچیدهی مهرهداران. او پیشنهاد داد که در فرگشت چشم انسان ممکن است چنین اندامهایی به عنوان حدواسط عمل کرده باشند.
اما این پرسش که ما چهگونه میبینیم، همچنان باقی است. گرچه داروین توانست بخش اعظم جهانیان را متقاعد کند که چشم مدرن میتواند به طور تدریجی از ساختاری بسیار سادهتر ساخته شده باشد اما او هرگز تلاش نکرد تا توضیح دهد که یک لکهی سادهی حساس به نور که از نظر او نقطهی شروع فرگشت چشم بود، در واقع چهگونه کار میکند. در بحث مربوط به چشم، داروین پاسخ به پرسش مربوط به مکانیسم نهایی شکلگیری چشم را با این عبارات کنار گذاشت: «وضعیت ما برای شناخت چهگونگی حساس شدن سلول عصبی نسبت به نور، چندان بهتر از وضعیتمان در خصوص نحوهی پیدایش خود حیات نیست.»[2]
او برای طفره رفتن از پاسخ به این سوال دلیل خیلی خوبی داشت: علم قرن نوزدهم هنوز آنقدر پیشرفت نکرده بود که به این نقطه برسد تا بخواهد به چنین موضوعی نزدیک شود. آن زمان پاسخ به این پرسش که چشم چهگونه کار میکند به سادگی امکان پذیر نبود. اکنون میدانیم این اتفاق زمانی رخ میدهد که فوتونهای نور به شبکیه برخورد کنند. در حقیقت، امکان پاسخ به هیچ پرسشی در مورد سازوکارهای زیربنایی حیات در آن زمان وجود نداشت: چهگونه ماهیچههای جانوران باعث حرکت میشوند؟ فتوسنتز چهگونه کار میکند؟ چهگونه از غذا انرژی استخراج میشود؟ بدن چهگونه با عفونت مقابله میکند؟ کسی چیزی نمیدانست.
کالوینیسم
اکنون به نظر میرسد این خصوصیت ذهن انسان باشد که وقتی دانش مربوط به سازوکارهای یک فرایند او را مقید نکرده باشد، تصور اینکه پدیدهای بتواند با چند گام ساده، از فقدان عملکرد به عملکرد برسد برایش ساده خواهد بود. یک مثال سرخوشانه در این مورد در مجموعهی کتابهای فکاهی مشهور کالوین[3] و هابز[4] ذکر شده است. کالوین کوچولو همیشه به همراه ببرش، هابز، به ماجراجویی میپردازد. آنها در جعبهای میپرند و در زمان به گذشته سفر میکنند، یا یک تفنگ اسباببازی اشعهای به دست میگیرند و خودشان را به شکل انواع حیوانات مختلف در میآورند یا بار دیگر از یک جعبه به عنوان همسانساز استفاده میکنند و کالوین کپیهای متعددی از خودش درست میکند تا با قدرتهای جهانی مانند مادرش یا معلمهایش مقابله کند. شاید برای بچهی کوچکی مانند کالوین تصور این که یک جعبه بتواند مانند هواپیما پرواز کند، یا چیزهایی از این دست، خیلی ساده باشد چون او نمیداند هواپیما چهگونه کار میکند.
یک مثال خوب از ساده فرض کردن تغییرات پیچیده در دنیای زیستشناسی باور به نظریهی «خلق خودبهخودی»[5] است. یکی از طرفداران اصلی این نظریه در اواسط قرن نوزدهم ارنست هکل[6] بود. او همچنین یکی از مروجان مشتاق نظریهی داروین هم بود. با توجه به این که در قرن نوزدهم دید محدودی به واسطهی میکروسکوپها در مورد سلول حاصل شده بود، هکل بر این عقیده بود که سلول یک «تودهی کوچک سادهی حاصل از ترکیب کربن و آلبومین است»[7] و تفاوت چندانی با یک ژلهی میکروسکوپی ندارد. بر همین اساس هکل معتقد بود که چنین حیات سادهای میتواند به سادگی از مواد غیرزنده تولید شود. در سال 1859، سال انتشار منشا انواع، یک کشتی اکتشافی به نام اچ.اماس سیکلوپس[8]، مقداری لجن با ظاهر عجیب و غریب از کف دریا بیرون کشید و نهایتا هکل به بررسی لجن پرداخت و به این نتیجه رسید که بسیار شبیه سلولهایی است که زیر میکروسکوپ دیده است. او در حالی که بسیار هیجان زده شده بود، توجه دوست صمیمی و مدافع بزرگ داروین، توماس هنری هاکسلی[9]، را معطوف این کشفش کرد. هاکسلی هم متقاعد شد که این همان لجن اولیهی پیشساز حیات است و آن را به احترام طرفدار برجستهی نظریهی شکلگیری خودبهخودی موجودات، بثیبیوس هاکلی[10] نام نهاد.
لجن موفق نشد رشد کند. سالها بعد با پیشرفت تکنیکهای زیستشیمیایی جدید و بهبود میکروسکوپها، پیچیدگی سلولها آشکار شد. معلوم شد که آن «تودهی ساده» شامل هزاران نوع مولکول آلی مختلف، پروتئینها، نوکلئیکاسیدها، بسیاری از ساختارهای زیرسلولی، ترکیبات خاص جهت فرایند تمایز و یک معماری بسیار پیچیده است. اگر اکنون با چشمانداز زمان خود به عقب نگاه کنیم، داستان بثیبیوس هاکلی به نظرمان احمقانه یا شاید حتی شرمآور باشد، اما نباید چنین باشد. هکل و هاکسلی کاملا طبیعی رفتار کردند، مانند کالوین، از آنجا که آنها از پیچیدگی سلولها آگاهی نداشتند، برایشان بسیار ساده بود که فرض کنند سلولها میتوانند از یک لجن ساده شکل بگیرند.
در طول تاریخ مثالهای بسیاری مشابه داستان هکل، هاکسلی و سلول وجود داشته است؛ جایی که یک تکه از یک معمای علمی خاص فراتر از درک آن زمانه بوده است. در علم یک اصطلاح جالب برای ماشینها یا ساختارها یا فرایندهایی که کاری را انجام میدهند اما سازوکار دقیقی که طی آن وظایفشان را انجام میدهند، ناشناخته است، وجود دارد. پدیدههای این چنینی را «جعبه سیاه» میخوانند. در دورهی داروین تمام زیستشناسی یک جعبه سیاه بود؛ نه فقط سلول یا چشم یا سیستم گوارشی یا ایمنی، بلکه تمام ساختارها و عملکردهای زیستشناسی. دلیلش هم این بود که هیچ کس نمیتوانست توضیحی ارایه دهد که فرایندهای زیستشناختی چهگونه به وقوع میپیوندند.
ارنست مایر[11]، مورخ و زیستشناس برجسته و نیروی هدایتکنندهای که پشت ترکیب نوداروینی ایستاده است، مینویسد:
هر انقلاب علمی باید همهی انواع جعبههای سیاه را بپذیرد، زیرا اگر بخواهد منتظر بماند تا تمام جعبههای سیاه باز شوند، هیچ وقت پیشرفتهای مفهومی نخواهد داشت.[12]
این یک حقیقت است اما در همان روزهای اولی که جعبههای سیاه باز شدند، به نظر رسید که علم و گاهی کل جهان تغییر کرد. زیستشناسی مطابق مدلی که داروین پیش رو نهاد، شاهد پیشرفتهای چشمگیری بود اما جعبههای سیاهی که داروین پذیرفته بود حالا در حال گشوده شدناند و دیدگاه ما به جهان بار دیگر متزلزل شده است.
پروتئینها
برای درک بنیانها مولکولی حیات، لازم است سر در بیاوریم که «پروتئینها» چهگونه کار میکنند. گرچه بسیاری از مردم به پروتئینها به چشم خوردنیهایی که بخش اعظم گروههای غذایی را تشکیل دادهاند، نگاه میکنند اما زمانی که پروتئینها را در بدن یک جانور یا گیاه خورده نشده و زنده در نظر میگیریم، میبینیم که در راستای مقاصد متفاوتی خدمت میکنند. پروتئینها ابزارآلات بافتهای زندهاند. آنها ساختارها را میسازند و واکنش شیمیایی لازم برای حیات را پیش میبرند. برای مثال، اولین مرحله از مراحل متعدد مورد نیاز برای تبدیل قند به انرژی زیستی قابل استفاده را پروتئینهایی به نام هگزوکیناز انجام میدهند. بخش اعظم اجزای سازندهی پوست را پروتئینی به نام کلاژن تشکیل میدهد. زمانی که نور به شبکیه برخورد میکند، ابتدا با پروتئینی به نام ردوپسین برهمکنش ایجاد میکند. همانطور که قابل مشاهده است، در همین مثالهای محدود هم میتوانیم ببینیم که عملکرد پروتئینها به طرز شگفتانگیزی متنوع است. با وجود این، به طورکلی یک پروتئین معین تنها یک یا چند عملکرد میتواند داشته باشد: ردوپسین نمیتواند پوست بسازد و کلاژن نمیتواند به طرزی کارآمد با نور واکنش دهد. بنابراین، یک سلول معمولی شامل هزاران هزار انواع پروتئین متفاوت است تا از پس وظایف متعدد لازم برای حیات بر آید؛ درست مانند یک کارگاه نجاری که شامل انواع مختلفی از ابزارآلات نجاری است.
این ابزارهای همهکاره چه شکلی هستند؟ ساختار پایهای پروتئینها تقریبا ساده است: اجزای سازندهی آنها که آمینواسید نامیده میشود، به صورت واحدهای مجزا به هم قلاب میشوند و تشکیل یک زنجیر میدهند. گرچه زنجیرهی پروتئینی میتواند از 50 تا 1000 آمینواسید داشته باشد اما این آمینواسیدها تنها میتوانند یکی از بیست آمینواسید معین باشند. از این منظر پروتئینها شبیه کلمات هستند. کلمات میتوانند هر طولی داشته باشند اما تنها میتوانند از 26 حرف مشخص ساخته شده باشند. از طرفی دیگر، یک پروتئین درون سلول مانند یک زنجیر شل و ول شناور نمیماند، بلکه تا میخورد و به یک ساختار بسیار دقیق در میآید که از پروتئینی به پروتئین دیگر متفاوت است. زمانی که همهی کارها انجام شد، دو توالی آمینواسیدی مختلف – دو پروتئین مختلف – میتوانند طوری خاص و متفاوت از هم تا بخورند که تفاوتشان در حد آچار فرانسه و ارهای مویی باشد و درست مانند ابزار و یراق آلات خانگی، اگر پروتئینی به طور قابل ملاحظهای از شکلش خارج شود، دیگر قادر نخواهد بود کارش را انجام دهد.
بینایی انسان
به طور کلی، این شبکههای پروتئینی هستند که فرایندهای زیستی را در سطح مولکولی به اجرا در میآورند، به طوری که هر کدام از اعضای این شبکهها وظیفهی خاصی را در زنجیرهای از وظایف برعهده دارد.
اجازه دهید بازگردیم به همان پرسشِ «ما چهگونه میبینیم؟». گرچه برای داروین رویداد اولیهی دیدن یک جعبه سیاه محسوب میشد، اما حالا با تلاشهای بسیاری از زیستشیمیدانان پاسخی برای این پرسش در دست است.[13]زمانی که نور به شبکیه برخورد میکند، یک مولکول آلی به نام 11-سیس-رتینال[14] فوتون نور را جذب میکند و ظرف چند پیکوثانیه دچار بازآرایی میشود و به ترنس-رتینال تبدیل میشود. تغییر شکل رتینال باعث تغییر شکلی قابل ملاحظه در پروتئین ردوپسین که پیوندی تنگاتنگ با آن دارد، میشود. در پی این تغییر شکل پروتئین، رفتار پروتئین هم از بسیاری جهات تغییر میکند. پروتئین دگرگون شده حالا میتواند با پروتئین دیگری که ترنسدوسین خوانده میشود، برهمکنش برقرار کند. قبل از تعامل ترنسدوسین با ردوپسین، این مولکول به یک مولکول کوچک آلی به نام GDP متصل است، اما وقتی که با ردوپسین پیوند برقرار میکند GDP خودش را از ترنسدوسین جدا میکند و مولکولی به نام GTP به ترنسدوسین میپیوندد. GTP و GDP اگرچه تا حد زیادی با یکدیگر خویشاوندند، اما تفاوتهایی اساسی با هم دارند.
آمدن GTP به جای GDP در مجموعهی ترنسدوسین-ردوپسین، رفتار این مجموعه را تغییر میدهد. GTP ترنسدوسین-ردوپسینی به پروتئینی در غشای داخلی سلول به نام فسفودیاستراز میپیوندد. با پیوستن ردوپسین و ملحقاتش به فسفودیاستراز، فسفودیاستراز توانایی تجزیهی شیمیایی مولکولی به نام cGMP را کسب میکند. ابتدا مولکولهای cGMP بسیاری درون سلول وجود دارند اما با فعال شدن فسفودیاستراز از غلظت cGMP کاسته میشود. فعال شدن فسفودیاستراز را میتوان به برداشتن در پوش وان حمام تشبیه کرد که باعث کاهش سطح آب میشود. پروتئین غشایی دیگری که با cGMP پیوند برقرار میکند، کانال یونی خوانده میشود که میتوانیم آن را یک مسیر اختصاصی برای تنظیم تعداد سدیمهای درون سلول در نظر بگیریم. کانال یونی در حالت عادی به سدیمها این امکان را میدهد که به درون سلول جاری شوند، در حالی که یک پروتئین دیگر به طور جداگانه با مصرف انرژی سدیمها را دوباره به خارج از سلول پمپ میکند. فعالیت دوگانهی کانال یونی و پروتئینهای پمپ، سطح یونهای سدیم سلول را در یک محدودهی باریک نگه میدارد. زمانی که غلظت cGMP بهواسطهی تجزیه شدنش به کمک فسفودیاستراز از مقدار معمولش کمتر میشود، بسیاری از کانالهای یونی بسته میشوند و در پی آن غلظت یونهای سدیم مثبت سلول هم کاهش مییابد. این رخداد منجر به بهم خوردن تعادل بار در دو سوی غشای سلولی میشود که در نهایت منجر به هدایت جریانی از عصب بینایی به مغز میشود. نتیجهی تمام این رخدادها و بعد از تفسیر پیام از سوی مغز، بینایی خواهد بود.
اگر زیستشیمی بینایی به همین فهرست واکنشهای بالا محدود میشد، سلول به سرعت از 11-سیس-رتینال و cGMP خالی میشد و به همین ترتیب تمام ذخیرهی سدیم داخل سلولی هم تخلیه میشد. بنابراین به سیستمی نیاز است که سیگنال تولید شده را محدود کند و سلول را به حالت اولیهاش بازگرداند. مکانیسمهای متعددی وجود دارند که این این عمل را انجام میدهند. به طور معمول، در تاریکی، کانال یونی علاوه بر یونهای سدیم به یونهای کلسیم هم اجازهی ورود به سلول میدهد؛ کلسیم با یک پروتئین دیگر به بیرون پمپ میشود تا غلظت آن درون سلول ثابت باشد. اگرچه با کاهش سطح غلظت cGMP، خاموش شدن کانالهای یونی و کاهش سطح غلظت یون سدیم، غلظت یون کلسیم هم کاهش مییابد. آنزیم فسفودیاستراز، که وظیفهی منهدم کردن cGMP را بر عهده دارد، با کاهش غلظت کلسیم، به شدت سرعت عملکردش کاهش مییابد. بهعلاوه، زمانی که غلظت کلسیم شروع رو به کاهش میرود، پروتئینی به نام گوآنیلات سیکلاز شروع به سنتز دوبارهی cGMP میکند. در همین حال، همزمان که همهی این رخدادها در حال وقوعاند، آنزیمی به نام ردوپسین کیناز تغییرات شیمیاییای در متاردوپسین II ایجاد میکند و یک گروه فسفات روی سوبسترا قرار میدهد. ردوپسین تغییر شکل یافته سپس به یک پروتئین دوگانهی دیگر به نام ارستین متصل میشود که جلوی ردوپسین را میگیرد تا ترنسدوسینهای بیشتری را فعال نکند. بنابراین، سلولها سازوکارهایی دارند که تقویت مستمر سیگنالی را که با یک فوتون آغاز شده بود محدود میکند.
ترنس رتینال در نهایت از مولکول ردوپسین کنده میشود و باید به 11-سیس-رتینال تبدیل شود و بار دیگر با پیوستن به اپسین، ردوپسین را میسازند تا چرخهی بینایی دیگری رقم بزنند. برای انجام این فرایند، ابتدا آنزیمی تغییرات شیمیاییای در ترنس-رتینال ایجاد میکند و سپس با افزودن دو هیدروژن آن را تبدیل به ترنسرتینول میکند. در مرحلهی بعد آنزیمی دیگر این مولکول را ایزومریزه کرده و به 11-سیس-تینول تبدیل میکند. در نهایت، یک آنزیم سوم اتمهای هیدروژنی را که پیشتر اضافه شده بودند حذف میکند تا 11-سیس-رتینال شکل بگیرد و چرخه کامل شود.
به سوی تبیین حیات
گرچه بسیاری از جزییات زیستشیمیایی بینایی در اینجا ذکر نشده است اما این نمای کلی که ملاحظه شد، نهایتا نشان میدهد که وقتی بنابر «تبیین» بینایی باشد، این چیزی است که باید ارایه شود. این سطحی از توضیح و تشریح است که علوم زیستی باید در نهایت هدف قرار دهند. برای این که بتوانیم بگوییم عملکردی را به طور کامل درک کردهایم، تمام مراحل مربوطه در فرایند باید روشن و مشخص شده باشند. مراحل مربوطه در فرایندهای زیستی نهایتا در سطوح مولکولی به وقوع میپیوندند، از همین رو توضیح پدیدههای زیستیای از قبیل بینایی یا گوارش یا ایمنی تنها در صورتی میتواند رضایتبخش باشد که توضیحی مولکولی از آن پدیده ارایه دهیم. حالا که جعبهی سیاه بینایی گشوده شده است، دیگر صرفا «توضیحات فرگشتیای» مانند آنچه داروین در قرن نوزدهم ارایه میداد یا آنچه مروجان فرگشت امروزه ارایه میدهند و تنها به ساختارهای آناتومیکی کل چشم استناد میکنند، به هیچ وجه کافی و رضایتبخش نیستند. آناتومی تا حدود زیادی بیربط است. گزارشهای فسیلی هم به همین ترتیب. دیگر اهمیتی ندارد که گزارشهای فسیلی با نظریهی فرگشت همسو است یا نیست، همانطور که دیگر اهمیتی ندارد فیزیک نیوتونی با تجربهی هر روزهی ما در زندگی همسو است. گزارشهای فسیلی در مورد این که تعاملات بین 11- سیس-رتینال با ردوپسین، ترنسدوسین و فسفودیاستراز چهگونه پله پله توسعه یافته و شکل گرفته است، چیزی برای گفتن ندارند. همان طور که الگوهای زیستجغرافیایی یا ژنتیک جمعیت یا توضیحاتی که نظریهی فرگشت در مورد اندامهای اولیه یا فراوانی گونهها میدهد، حرفی برای زدن ندارند.
داروین در قرن نوزدهم گفت: «وضعیت ما برای شناخت چهگونگی حساس شدن سلول عصبی نسبت به نور، چندان بهتر از وضعیتمان در برابر نحوهی پیدایش خود حیات نیست.» اما هر دو پدیده نظر زیستشیمیدانان مدرن را به سوی خود جذب کردهاند. داستان سکتهی خفیفی که تحقیقات مربوط به خاستگاه حیات دچارش شد، بسیار هیجانانگیز است اما متاسفانه در این مقاله نمیگنجد. از همین رو تنها به بیان این نکته بسنده میکنم که در حال حاضر رشتهی مطالعات خاستگاهحیات در ترکیبی بدآهنگ از مدلهای متناقض غرق شده است که همگی نامتقاعدکننده و به طرزی جدی غیرکاملاند و هیچ سازگاریای با مدل رقابتی ندارند. حتی بیشتر زیستشناسان فرگشتی در خلوت خودشان اذعان میکنند که علم هیچ توضیحی برای منشا حیات ندارد.
هدف این مقاله این است که نشان دهد تحقیقات مربوط به خاستگاه حیات با همان مشکلهایی دست و پنجه نرم میکند که تلاشها برای توضیح منشا سیستمهای پیچیده با آنها دست به گریبان است. زیستشیمی جهانی مولکولی را آشکار کرده است که در برابر توضیحاتی که سالیانی دراز در سطوح کلی موجودات اعمال میشدند، با قدرت مقاومت میکند. هیچکدام از جعبههای سیاه داروین را -چه منشا حیات، چه منشا بینایی یا هر سیستم زیستشیمیایی دیگر را- تاکنون نظریهی خود او تبیین نکردهاند.
پیچیدگی کاهشناپذیر
داروین در منشا انواع مینویسد: «اگر اندامی پیدا میشد که از طریق تغییرات متعدد، متوالی و سبک پدید نیامده باشد، بطلان فرضیهی من به اثبات میرسید.»[15] سیستمی که با معیار داروین همخوانی داشته باشد، در حقیقت پیچیدگیکاهشناپذیر[16] را به نمایش میگذارد. منظورم از پیچیدگیکاهشناپذیر نظام واحدی است که از تعامل چندین بخش ساخته شده است. این بخشها برای یک عملکرد اصلی با هم همکاری میکنند و حذف هر کدام از این بخشها منجر به از کار افتادن عملکرد موثر سیستم میشود. یک سیستم پیچیدهیکاهشناپذیر نمیتواند در اثر «تغییرات تدریجی، سبک و متوالی» سیستم قبلی ساخته شود. چرا که طبق تعریف پیچیدگیکاهشناپذیر، هر سیستمی قبل از سیستم فعلی ناکارآمد و بدون عملکرد است. از آنجا که انتخاب طبیعی نیاز به عملکردی دارد تا بتواند انتخابش کند، یک سیستم پیچیدهیکاهشناپذیر زیستی (اگر همچین چیزی وجود داشته باشد) باید به صورت یک واحد یکپارچه ظهور پیدا کرده باشد تا انتخاب طبیعی چیزی داشته باشد که بتواند روی آن عملی انجام دهد. تقریبا همگان پذیرفتهاند که چنین رویداد ناگهانیای با آن فرایند تدریجیای که داروین تصور میکرد، سر سازگاری ندارد. از این نظر، اگرچه «پیچیدگیکاهشناپذیر» تنها یک عبارت است، اما قدرتش در مفهومش نهفته است. حال باید بپرسیم که آیا در واقعیت هم چیزی با پیچیدگی کاهشناپذیر وجود دارد و اگر وجود دارد، چنین چیزی در سیستمهای زیستی هم وجود دارد؟
یک تله موش معمولی را در نظر بگیرید (شکل 1). تله موشی که خانوادهی من در خانه استفاده میکنند تا به حساب جوندگان مزاحم برسند، از چند بخش تشکیل شده است: 1) یک صفحهی چوبی مسطح که به عنوان زمینه و تکیهگاه عمل میکند؛ 2) یک چکش فلزی که عمل اصلی کشتن موش را انجام میدهد؛ 3) یک فنر سیمی با انتهاهای ادامه یافته برای این که در زمان عمل تله به بستر و چکش نیرو وارد کند؛ 4) یک چفت بسیار حساس که با اعمال کوچکترین فشار آزاد میشود؛ 5) یک میلهی فلزی که وقتی تله در حالت آماده باش است، چکش را عقب نگه دارد و به چفت متصل میشود. همچنین پیچ و مهرههایی وجود دارد که کل سیستم را کنار هم نگه میدارند.
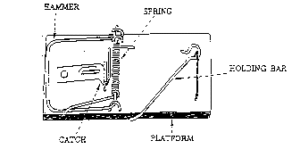
شکل 1. یک تلهموش خانگی. اجزای عملکننده علامتگذاری شدهاند. اگر هر کدام از اجزا از دست برود، تله از کار میافتد.
اگر هر کدام از اجزای تلهموش (تکیهگاه، چکش، فنر، چفت و میلهی نگهدارنده) حذف شود، تله دیگر کار نمیکند. به عبارت دیگر، یک تلهموش سادهی کوچک تا زمانی که همهی اجزایش در کنار هم قرار نگرفته باشند، هیچ تواناییای ندارد.
از آنجا که تلهموش الزاما از بخشهای مختلف تشکیل شده است، پس نوعی پیچیدگیکاهشناپذیر است. در نتیجه، پیچیدگیکاهشناپذیر در واقعیت وجود دارد.
ماشینهای مولکولی
حالا ببینیم آیا سیستم زیستشیمیایی پیچیدهیکاهشناپذیر هم وجود دارد؟ بله، مشخص شده است که تعداد بسیاری از آنها وجود دارد.
پیشتر در مورد پروتئینها صحبت کردیم. در بسیاری از ساختارهای زیستی، پروتئینها صرفا اجزای سازندهی ماشینهای مولکولی بزرگترند. مانند لامپ تصویر، سیمها و پیچ و مهرههایی که یک دستگاه تلویزیون را میسازند، بسیاری از پروتئینها اجزای ساختارهایی هستند که تنها زمانی کار میکنند که تقریبا تمام اجزای سازنده کنار هم قرار داشته باشد. یک مثال خوب در این زمینه مژک است.[17]
مژکها اندامهای مومانندی هستند که در سطح بسیاری از سلولهای جانوری یا سلولهای گیاهان پستتر وجود دارند و مایعات را روی سطح سلول جابهجا میکنند یا با عملی مانند «پارو زدن» تک سلولیها را میان یک سیال به جلو میرانند. در انسان، برای مثال سلولهای اپیتلیالی که سطح محفظهی تنفسی را پوشاندهاند، هر کدام حدود 200 مژک دارند که به طور هماهنگ با هم ضربه میزنند تا مخاط را به سوی گلو برانند تا دفع شود. یک مژک از یک دستهی تارهای پوشیده شده با غشا به نام آکسونم تشکیل شده است. یک آکسونم شامل یک حلقه از نه جفت میکروتوبول است که اطراف دو میکروتوبول منفرد مرکزی را احاطه کرده است. هر جفت بیرونی از حلقه از سیزده رشته (ریزرشتهی A) در هم تنیده با ده رشته (ریز رشتهیB ) تشکیل شده است. رشتههای میکروتوبولی آمیختهای از دو پروتئین به نامهای توبولین آلفا و بتا است. یازده میکروتوبول تشکیل یک آکسونم میدهند که ازطریق سه نوع پیوند دهنده کنار یکدیگر نگهداری میشوند. ریزرشتههای A با میلههای شعاعی به میکروتوبولهای مرکزی متصل میشوند. جفتهای همجوار بیرونیتر با پیونددهندههایی به هم متصل میشوند که از پروتئینهایی بسیار کشسان به نام نکسین تشکیل شدهاند؛ و میکروتوبولهای مرکزی با پلهای متصلکننده به یکدیگر متصل شدهاند. در نهایت، هر ریزرشته A دارای دو بازو است، بازوی داخلی و بازوی خارجی که هر دو حاوی پروتئین داینئین هستند.
اما یک مژک چهگونه کار میکند؟ آزمایشها نشان دادهاند که حرکت مژک در نتیجهی «راه رفتن» بازوهای داینئین یک میکروتوبول روی ریزرشتهی B میکروتوبول دومی و با مصرف انرژی شیمیایی حاصل میشود. بدین ترتیب دو میکروتوبول روی هم سر میخورند و میگذرند (شکل 2) با وجود این، پیوندهای عرضی پروتئینی بین میکروتوبولها در یک مژک سالم نمیگذارد که میکروتوبولهای همسایه بیش از یک فاصلهی کوتاه از کنار یکدیگر گذر کنند. بدین ترتیب، این پیوندهای عرضی حرکت کشویی داینئینها را به حرکت خمشی کل آکسونم تبدیل میکنند.
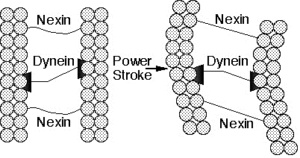
شکل 2. طراحی شماتیک از بخشی از یک مژک. حرکات موتور پروتئینی داینئین، متصل به یک میکروتوبول، روی ریزرشتهی B میکروتوبول همسایه باعث میشود که این رشتهها روی یکدیگر بلغزند. پروتئین ارتجاعی نکسین، حرکت لغزشی را به حرکت خمشی تبدیل میکند.
اکنون اجازه دهید نحوهی کار مژک را مرور کنیم و ببینیم این موضوع بر چه چیزی دلالت دارد. مژکها از حداقل نیم دوجین پروتئین تشکیل شدهاند: آلفا توبولین، بتا توبولین، داینئین، نکسین، پروتئین میلهای و یک پروتئین پل مرکزی. همهی اینها با هم ترکیب میشوند تا یک وظیفهی معین، یعنی حرکت مژکی را رقم بزنند و حضور همهی این پروتئینها برای عملکرد داشتن مژک الزامی است. اگر توبولینها غایب باشند، آنگاه رشتههایی وجود نخواهد داشت که بخواهد روی هم سر بخورند. در صورت نبود داینئین، آنگاه مژک صلب و بیحرکت باقی میماند. اگر نکسین یا سایر پروتئینهای متصل کننده وجود نداشته باشند، آنگاه وقتی رشتهها روی هم میلغزند آکسونم از هم جدا میشود.
آنچه در مژک میبینیم، صرفا یک پیچیدگی عمیق نیست، بلکه یک پیچیدگیکاهشناپذیر در سطح مولکولی است. در خاطر داشته باشید که منظور ما از «پیچیدگیکاهشناپذیر» دستگاهی است که برای کارکرد خود نیازمند چندین جزء مجزا است. تلهموش من باید یک تکیهگاه، چکش، فنر، چفت و میلهی نگهدارنده داشته باشد و همگی با هم کار کنند تا تلهموش عملکرد داشته باشد. به طور مشابه، مژک هم باید رشتههای لغزنده، پروتئینهای متصل کننده و موتور پروتئینی داشته باشد تا عملش امکانپذیر شود. در غیاب هر کدام از این اجزا، کل این سیستم بیاثر میشود.
اجزای مژک مولکولهای منفرد هستند. این بدان معنا است که دیگر جعبه سیاهی در کار نیست که دست به دامانش شویم. پیچیدگی مژک، پیچیدگیای نهایی و بنیادین است. همانطور که دانشمندان زمانی که شروع به یادگیری پیچیدگیهای موجود در سلول کردند، پی بردند تفکرشان بسیار احمقانه بوده که فکر میکردند حیات به طور خود به خودی و طی یک یا چند گام از لجن اقیانوس شکل میگیرد، حالا هم ما میدانیم که پیچیدگی مژک با یک یا چند گام ساده بدست نمیآید. از آنجا که پیچیدگیکاهشناپذیر مژک نمیتواند عملکرد پیشین داشته باشد، نمیتواند از طریق انتخاب طبیعی شکل گرفته باشد، زیرا انتخاب طبیعی به یک پیوستگی عملکردی نیاز دارد تا بتواند کار کند. وقتی عملکردی برای انتخابکردن نباشد، انتخاب طبیعی ناتوان میشود. ما میتوانیم یک گام فراتر بگذاریم و بگوییم که اگر انتخاب طبیعی نمیتواند مژک بسازد، پس مژک طراحی شده است.
مطالعهی «فرگشت مولکولی»
مثالهای دیگر از پیچیدگیکاهشناپذیر فراواناند، از جمله جنبههای انتقال پروتئین، انعقاد خون، DNA بستهی حلقوی، انتقال الکترون، تاژک باکتری، تلومرها، فتوسنتز، تنظیم رونویسی و نمونههای دیگر. مثالهای پیچیدگیکاهشناپذیر را تقریبا در هر صفحه از یک کتاب درسی زیستشیمی میتوانید پیدا کنید. اما اگر این چیزها را نمیتوان با فرگشت داروین توضیح داد، پس جامعهی علمی با چه دیدی به این پدیدهها در چهل سال گذشته نگریسته است؟ یک جای مناسب برای یافتن پاسخ این پرسش مجلهی جورنال آو مولکولار اوولوشن[18] است. این مجله است کارش را اختصاصا برای بررسی این موضوع شروع کرد که فرگشت چهگونه در سطح مولکولی اتفاق میافتد. این مجله استانداردهای علمی بالایی دارد و ویراستاران آن چهرههای برجستهی این حوزه هستند .در یکی از شمارههای اخیر این نشریه، یازده مقاله منتشر شده بود که تقریبا همهی آنها در مورد آنالیز پروتئینها یا توالیهای DNA بودند. در هیچ کدام از این مقالهها در مورد مدلهای دقیق بینابینی در طول تکامل ساختارهای زیستشیمیایی پیچیده سخنی به میان نیامده است. این مجله در ده سال گذشته 886 مقاله منتشر کرده که در این میان، 95 مقاله به بحث در مورد سنتز شیمیایی مولکولهای مورد نیاز برای پیدایش حیات پرداخته است، 44 مقاله مدلهای ریاضیاتی برای بهبود آنالیزهای توالییابی را ارایه دادهاند، 20 مقاله به پیامدهای تکاملی ساختارهای فعلی پرداختهاند و 719 مورد در زمینهی تجزیه و تحلیل پروتئینها یا توالیهای پلینوکلئوتیدی بودهاند. هیچ مقالهای نبود که مدلی دقیق برای حالتهای بینابینی در توسعهی ساختارهای پیچیدهی مولکولهای زیستی ارایه دهد. این موضوع مختص مجلهی جورنال آو مولکولار اوولوشن نیست. در هیچ کدام از نشریات جورنال آو مولکولار بایولوژی[19]، ساینس[20]، نیچر[21] و پروسیدینگ آو ذ نشنال آکادمی آو ساینس[22] هیچ مقالهای پیدا نمیکنید که سخنی از مدلی دقیق برای حالتهای بینابینی در توسعهی ساختارهای مولکولهای زیستی پیچیده به میان آورده باشد. تا جایی که میدانم در هیچ مجلهی دیگری هم به حالتهای بینابینی پرداخته نشده است.
مقایسهی توالیها به طرز کلافهکنندهای بر نوشتههایی در مورد فرگشت مولکولی غالب است اما مقایسهی توالیها همانقدر میتواند به توجیه نحوهی رشد و شکلگیری سیستمهای پیچیدهی زیستشیمیایی کمک کند که قیاس بین چشمهای ساده و پیچیده، به داروین کمک کرد تا بتواند بگوید که بینایی چهگونه اتفاق میافتد. بنابراین، در این حوزه علم به طور کامل سکوت اختیار کرده است. این بدان معنا است که وقتی استنباط میکنیم سیستمهای زیستشیمیایی پیچیده طراحی شدهاند، با هیچ نتیجهی آزمایشی یا مطالعهی نظری در تضاد نیستیم و آن را نقض نکردهایم. در واقع نیازی برای به تردید افکندن هیچ آزمایشی نیست بلکه باید تفاسیر همهی آزمایشها را بار دیگر مورد بررسی قرار داد.
نتیجهگیری
اغلب گفته میشود علم باید از نتیجهگیریهایی که رنگ و بوی فراطبیعی دارند پرهیز کند. اما به نظرم این هم یک منطق و هم یک علم بد است. علم بازی نیست که قوانین سفت و سخت و مستبدانه تعیین کنند چه تبیینهایی مجازند و کدامها مجاز نیستند. بلکه علم تلاشی است برای آنکه بیانی صحیح در مورد واقعیت فیزیکی ارایه شود. تنها شصت سال پیش بود که برای اولین بار انبساط جهان مشاهده شد. این واقعیت بلافاصله پیشنهاد یک رویداد خارقالعاده را مطرح کرد، این که در گذشتههای بسیار دور کل جهان از یک ذرهی بینهایت کوچک شروع به انبساط کرده است. برای بسیاری از افراد این استنباط رنگ و بوی یک رویداد فراطبیعی داشت: خلقت و آغاز عالم. فیزیکدان برجستهی انگلیسی، آرتور استنلی ادینگتون[23]، احتمالا با لحنی آمیخته با انزجار برای بسیاری از فیزیکدانان از این عقیده سخن گفته است:
از نظر فلسفی، یک شروع ناگهانی برای نظم فعلی طبیعت، از نظر من و فکر میکنم از نظر بسیاری، ایدهای متناقض و مطرود است و حتی آنهایی که از این ایده به عنوان اثباتی بر مداخلهی یک خالق استقبال میکنند، احتمالا در نظر خواهند گرفت که یک واقعهی منفرد در دورانی بسیار دور، واقعا آن نوع رابطهای بین خدا و جهانش نیست که احساس رضایت و خرسندی به ذهن متبادر کند.[24]
به هر ترتیب، فرضیه مهبانگ از سوی فیزیک پذیرفته شد و طی سالهای گذشته ثابت شده است که یک الگوی بسیار ثمربخش است. نکته اینجا است که حتی در جایی که برخی معتقد بودند مدل ارایه شده با مذهب دست به یکی کرده است، باز هم فیزیک از دادههایی که پیش رویش قرار داشت پیروی کرد. در این روزها که زیستشیمی مثالهایی از سیستمهای مولکولی ارایه میکند که به طرز شگفتانگیزی پیچیدهاند، سیستمهایی که از هرگونه تلاش برای توضیح نحوهی شکلگیری حتی یکی از آنها به سردی استقبال میشود، ما باید از فیزیک درس بگیریم. نتیجهگیری طراحی هم طبیعتا از دادهها سرچشمه میگیرد و ما نباید از آن شانه خالی کنیم بلکه باید آن رادر آغوش بکشیم و روی آن کار کنیم.
به عنوان نتیجهگیری، مهم است که درک کنیم ما براساس ندانستههایمان به طراحی استنباط نمیکنیم، بلکه این کار را از روی دانستههایمان انجام میدهیم. ما برای توجیه یک جعبهی سیاه به طراحی استنباط نمیکنیم، بلکه براساس یک جعبهی باز این کار را میکنیم. اگر فردی از یک فرهنگ بدوی یک اتومبیل ببیند ممکن است تصور کند که باد دارد آن را به حرکت در میآورد یا یک بز کوهی در زیر ماشین پنهان شده است، اما وقتی کاپوت را باز میکند و موتور را میبیند، بلافاصله متوجه میشود که آن خودرو طراحی شده است. به همین ترتیب زیستشیمی سلول را باز کرده است تا بررسی کند چه چیزی آن را به عمل در میآورد و میبینیم که این نیز طراحی شده است.
وقتی براساس مشاهدات علمی انجام شده، کشف شد که بسیاری از ویژگیهای زیستی را میتوان با استناد به اصول ظریف انتخاب طبیعی توجیه کرد، برای مردم قرن نوزدهم شوک بزرگی بود. برای ما هم در قرن بیستم بسیار تکاندهنده است اگر براساس مشاهدات علمی انجام شده، کشف کنیم که سازوکارهای بنیادین حیات را نمیتوانیم به انتخاب طبیعی نسبت دهیم و از همین رو طراحی شدهاند. اما بهترین کاری که میتوانیم بکنیم این است که با این بهتمان کنار بیاییم و ادامه دهیم. نظریهی فرگشت هدایت نشده حالا دیگر مرده است، اما کار علم همچنان ادامه دارد.
منبع:
http://www.arn.org/docs/behe/mb_mm92496.htm
این مقاله در اصل در تابستان سال 1994 در جلسهی انجمن سی. اس. لویس دانشگاه کمبریج ارایه شد.
[1] The Origin of Species
[2] Darwin, Charles (1872) Origin of Species 6th ed (1988), p.151, New York University Press, New York.
[3]Calvin
[4] Hobbes
[5] spontaneous generation
[6] Ernst Haeckel
[7] Farley, John (1979) The Spontaneous Generation Controversy from Descartes to Oparin, 2nd ed, p.73, The Johns Hopkins University Press, Baltimore
[8] H.M.S. Cyclops
[9] Thomas Henry Huxley
[10] Bathybius Haeckelii
[11] Ernst Mayr
[12] Mayr, Ernst (1991) One Long Argument, p. 146, Harvard University Press, Cambridge
[13] Devlin, Thomas M. (1992) Textbook of Biochemistry, pp.938954, WileyLiss, New York
[14] 11-cis-retinal
[15] Darwin, p.154.
[16] Irreducible Complexity
[17] Voet, D. & Voet, J.G. (1990) Biochemistry, pp.11321139, John Wiley & Sons, New York.
[18] Journal of Molecular Evolution
[19] Journal of Molecular Biology
[20] Science
[21] Nature
[22] Proceedings of the National Academy of Science
[23] Arthur Stanley Eddington
[24] Cited in Jaki, Stanley L. (1980) Cosmos and Creator, pp.56, Gateway Editions, Chicago



