
تابع نمایی حیات: حیات در بسیاری از سطوح طراحی هوشمند را به نمایش میگذاردجاناتان ولز/ ترجمه: امیرحسین لطیفی

ژوئن 2018
سلول انسان شامل دو مجموعه DNA است که هر کدام در بردارندهی سه میلیارد زیرواحد به نام «نوکلئوتید» هستند. چهار نوع مختلف نوکلئوتید وجود دارد و میتوانند به بسیاری از روشهای مختلف آراسته شوند، بنابراین DNA تا حد زیادی پیچیده است. با وجود این، بخش زیادی از DNA ما باید به نحو بسیار خاص و مشخصی آرایش یابد که بتواند اطلاعات لازم را برای ساخت RNAها و پروتئینها فراهم آورد. ریاضیدان برجسته، ویلیام دمبسکی[1]، این پدیده را «اطلاعات پیچیدهی مشخص» مینامد.[1] پیچیدگی (مانند آنچه که در تودهای از برگهای پاییزی مشاهده میکنیم) میتواند به طور خودبهخودی از یک فرایند طبیعی هدایت نشده پدیدار شود، اما اطلاعات پیچیدهی مشخص نمیتوانند این چنین باشند. تنها منبع شناخته شده برای اطلاعات پیچیدهی مشخص یک ذهن هوشمند است که بتواند هدفی را مجسم کند و همه چیز را برای دست یافتن به آن هدف -که در اینجا منظور یک سلول زنده است- سازماندهی کند. استفان مِیر[2] فیلسوف علم در سال 1998 نوشت «از آن جا که ذهن یا طراحی هوشمند عاملی ضروری برای یک سیستم غنی از اطلاعات است، توالیهای نوکلئوتیدی که در DNA به نحوی مشخص آرایش یافتهاند – اطلاعات کدگذاری شده – بر عمل پیشین یک ذهن هوشمند دلالت دارند حتی اگر چنین عامل ذهنیای به طور مستقیم قابل مشاهده نباشد.»[2]
نیاز به اطلاعات مکانی
بنابراین، DNA اطلاعات زیستی را حمل میکند و این اطلاعات به طراحی منتهی میشوند. اما DNA تنها حامل اطلاعات در سلول زنده نیست. زمانی که DNA به RNA رونویسی میشود، بیشتر آن RNAها پیش از آن که بتوانند کارکرد مناسبی داشته باشد، باید به مناطق خاصی در سلول منتقل شوند. برخی RNAها با توالیهایی نشانهگذاری شدهاند که «کد پستی» خواند میشوند. این توالیها آدرسهایی را که RNAها باید به آنجا منتقل شوند، تعیین میکنند؛ مانند کد پستی روی نامهای که درون صندوق پست میاندازید. با وجود این، کدپستی یک RNA در صورتی که با یک آدرس از پیش موجود مطابقت نداشته باشد کاملا بیمعنا خواهد بود. مانند آدرسهای جغرافیایی در یک سیستم پستی، مقاصد سلولی RNAها و پروتئینها پیش از ارسال باید مستقل از کد پستیشان تعیین شده باشد. بسیاری از این مقصدها را مولکولهای قرار گرفته در غشای سلولی تعیین میکنند. دانشمندان ابتدا فکر میکردند که پروتئینها مانند قایقهای شناور در دریا میتوانند آزادانه در غشا پراکنده شوند اما امروزه میدانیم که بسیاری از پروتئینهای غشایی در الگوهایی غیرتصادفی سازمان یافتهاند که تا حد زیادی آنها را ثابت میکند. چنین الگوهایی اطلاعات مکانیای برای سلول فراهم میکنند که فراتر از اطلاعات موجود در DNA است.
یک تخم قورباغه بعد از لقاح به نحوی غیرقابل رویت به نواحیای تقسیم میشود که صرفا RNAهایی که در ناحیهای از غشای سلولی به نام «کورتکس» قرار دارند آنها را شناسایی میکنند. در شکل زیر چهار ناحیه با چهار رنگ مختلف مشخص شدهاند. بعد از این که هسته مضاعف شد تخم لقاح یافته به دو سلول دختری تقسیم میشود. اگر صفحهی تقسیم مطابق A باشد هر سلول دختر نه فقط هسته بلکه سهمی از اطلاعات قشری هر چهار ناحیه را به ارث میبرد. اما اگر صفحهی تقسیم مطابق B یا C باشد، سلولهای دختری مجموعهی کاملی از اطلاعات قشری به ارث نمیبرد و حتی اگر هر کدام اطلاعات DNA لازم را در برداشته باشند، باز هم تکوینشان متوقف میشود.
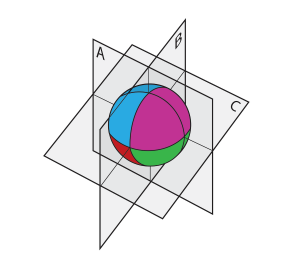
شکل 1. نواحی اطلاعات مکانی در یک تخم لقاح یافتهی قورباغه
حاملان اطلاعات مکانی دیگر
تفاوتهای ناحیهای در سلولها و جنین میتوانند به روشهای دیگری علاوه بر جایگذاری RNAها در کورتکس معین شوند. دو روش از این روشها به طور مفصل مورد مطالعه قرار گرفتهاند: «کد قندی» و «کد بیوالکتریکی».
بیشتر پروتئینهای سلول زنده –از جمله پروتئینهای غشایی– با پیوندهای شیمیایی به کربوهیدراتهایی به نام گلیکان (از واژهای یونانی به معنای «شیرین») متصلاند. این نوکلئوتیدها در DNA در مولکولی خطی به صورت پشت به پشت به هم متصلاند و بنابراین توالی اطلاعات DNA یک بُعدی است. در سلولهای زنده، زیرواحدها در پروتئینها هم (با برخی استثناها) به زنجیرهای خطی به هم متصلاند اما گلیکانها میتوانند به صورت مجموعههای سه بُعدی به هم متصل شوند، بنابراین ظرفیت حمل اطلاعات آنها چند ده برابر DNA و پروتئینها است.[4]
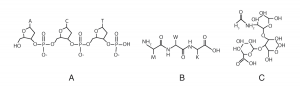
شکل 2. سه زیرواحدی که در A) DNA، B) پروتئین و C) گلیکان به هم متصل میشوند.
اطلاعاتی را که گلیکانها حمل میکنند «کد قندی» مینامند.[4] کد قندی با پروتئینهایی به نام لکتین تفسیر میشوند. این پروتئینها ساختارهای سهبُعدی خاص مولکولهای گلیکان را شناسایی میکنند. گلیکانها و لکتینها نقشی اساسی در ارتباط بین سلولها و کمک به حرکت سلولها در جنین در حال تکوین دارند. آزمایشها نشان دادهاند که الگوهای غشایی گلیکانها طی تکوین جنین تغییر میکنند.[5] علاوه بر کدها قندی، احتمالا همهی سلولهای زنده (نه فقط سلولهای عصبی و عضلانی) در اطراف غشایشان میدانهای الکتریکی میسازند. آنها این کار را با پمپ کردن یونهای باردار از میان کانالهای درون غشاهایشان «میدانهای بیوالکتریکی» میسازند. الگوی کانالهای غشایی میدانهای بیوالکتریکی را تعیین میکنند و فرم این میدانها در طول تکوین جنین تغییر میکنند.
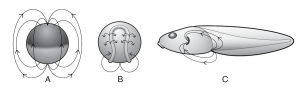
شکل 3. برخی از میدانهای الکتریکی (نشان داده شده با پیکان) در جنین در حال تکوین قورباغه
میدانهای بیوالکتریکی با رویدادهای تکوینی مهم در هم تنیدهاند. برای مثال در جنینهای قورباغه، جریانهای یونی بزرگ در مناطقی از اندامهای جلویی، خیلی قبل از این که این اندامها واقعا ظاهر شوند، تکوین مییابند.
بسیاری از آزمایشهایی که از سال 1980 انجام شده است، تایید کردهاند که اختلال در میدانهای بیوالکتریکی باعث اختلال در تکوین میشود. برای مثال، جنین قورباغه به طور معمول یک میدان الکتریکی در جهت سر به دم ایجاد میکند. اگر یک میدان مصنوعی با همان بزرگی در جهت مخالف اعمال شود یا اگر کانالهای یونی که میدانها را میسازند مسدود شوند، نتیجه تکوین غیرعادی سر و چشم خواهد بود. یعنی در نقاطی که به طور معمول چشمها تشکیل میشوند نسبت به بافت احاطهکننده تغییرات شدیدی را متحمل شده بود و در برخی موارد چشم در جای دیگری از بدن بچه قورباغه، حتی در دم آن، تکوین یافته بود.
میدانهای الکتریکی چهگونه تکوین را تحت تاثیر قرار میدهند؟ در دههی 1980 زیستشناسان سلولهای جنینی را در معرض میدانهای الکتریکی مصنوعیای با شدتی برابر با میدانهایی که سلولها در حالت طبیعی تولید میکنند، قرار دادند. برخی از انواع سلولها به سمت قطبهای مثبت مهاجرت کردند در حالی که سایر انواع به قطبهای منفی گسیل یافتند. این اتفاق این موضوع را پیشنهاد میکند که میدانهای الکتریکی، تکوین جنین را با هدایت جنبشهای سلولی تحت تاثیر قرار میدهند.
در سال 1995 دو زیستشناس به نامهای ریوی شی[3] و ریچارد بورگنز[4] به این نتیجه رسیدند که میدانهای بیوالکتریکی «احتمالا یک دستگاه مختصات سه بُعدی فراهم میکنند» که به تعیین اشکال در جنینها کمک میکند.[6] در سال 2013، زیستشناسان دیگری با نام آیسان تسنگ[5] و مایکل لوین[6] نوشتند که چنین میدانهایی احتمالا «قالبهای شکلی» فراهم میکنند و این که درک کامل از نحوهی تکوین جنین احتمالا مستلزم شکستن «کد بیوالکتریکی» است.[7]
کد غشا
بنابراین RNAهای متمرکز شده در کورتکس، الگوهای گلیکانهای روی غشا و میدانهای الکتریکی ساخته شده به وسیلهی کانالهای یونی در غشا همگی حامل اطلاعات مکانی هستند. هر چند مولکولهای منفرد ممکن است به وسیلهی DNA تعیین شوند اما الگوهای سهبُعدی آنها این چنین نیستند. در مجموع، این الگوها تشکیل یک «کد غشایی» میدهند که مستقل از توالیهای DNA است. در سال 1983 زیستشناس برجسته رابرت پویتون[7] پیشنهاد کرد که غشاهای زیستی حامل «خاطرات مکانی» هستند که واحدهای آنها پروتئینهای دارای استقرار فضایی معین است. پویتون نوشت: «درک این که خاطرهی ژنتیکی یکبُعدی و در امتداد یک مولکول DNA است در حالی که خاطرهی مکانی احتمالا در امتداد سطح غشا دو بُعدی و درون فضای داخلی سلول سهبُعدی است، بسیار محتمل است و نشان میدهد که خاطرهی مکانی ممکن است بسیار پیچیدهتر و متنوعتر از خاطرهی ژنتیکی باشد.»[8]
در سال 2004 توماس کاوالیه اسمیت[8] زیست شناس نوشت این ایده که DNA دربردارندهی همهی اطلاعات مورد نیاز برای ساخت یک موجود است «کاملا اشتباه است». طبق نظر کاوالیه اسمیت، غشاها «یک سطح شیمیایی دو بُعدی معین با روابط توپولوژیکی حفاظتشدهی دو جانبه در سه بُعد مکان دارند» که «نقشی کلیدی در مکانیسمهایی که در تبدیل اطلاعات DNA به اشکال سه بُعدی سلول منفرد و موجودات پرسلولی دارند، بازی میکنند. تکوین جانوران یک موجود پرسلولی سه بُعدی میسازد و این کار با اطلاعات خطی DNA آغاز نمیشود . . . بلکه شروع آن همواره از یک موجود سه بُعدی تک سلولی بسیار پیچیدهی از قبل موجود یعنی تخم لقاح یافته است.»[9]
بنابراین، کدها غشایی حامل اطلاعات زیستی اساسی هستند که مستقل از اطلاعات موجود در توالی DNA عمل میکنند. با این حال اغلب میشنویم که تکوین جنین را یک برنامهی موجود در DNA هدایت میکند. چرا؟
آن سوی DNA
جیمز واتسون[9] و فرانسیس کریک[10] برندگان جایزهی نوبل، ساختار مولکولی DNA را در سال 1954 کشف کردند. این رویداد به نظر میرسید که یک بنیان مولکولی نه فقط برای وراثت بلکه همچنین برای تکوین جنین فراهم کرده باشد. سلولها قبل از تقسیمشدن، DNAشان را همانندسازی میکنند و (معمولا) یک مجموعهی کامل از توالی DNAشان را به زادگانشان منتقل میکنند. سلولها سپس از توالیهای DNA به عنوان الگو برای رونویسی RNAها استفاده میکنند و برخی از این RNAها سپس به پروتئین ترجمه میشوند.
در میانهی قرن بیستم زیستشناسی تحت سلطهی نوداروینیسم قرار داشت، یک سسیستم فکری که فرگشت و ژنتیک را در هم آمیخت و تنوعات جدید را به جهشهای ژنتیکی نسبت داد. پیشفرض زیربنایی نوداروینیسم این است که فرگشت و تکوین کاملا ناشی از فرایندهای مادی هدایت نشده است. پس از سال 1953، این فرض مادهگرایانه به این دیدگاه منتج شد که «DNA، RNA را میسازد و آن هم پروتئین را میسازد و پروتئین هم ما را میسازد.» این دیدگاه قاعدهی اصلی زیستشناسی سلولی و مولکولی خوانده میشود.
در سال 1970 زیستشناس مولکولی (و ماتریالیست) ژاک مونود[11] گفت که با قاعدهی اصلی «و درک اساس فیزیکی تصادف در جهش که زیستشناسی مولکولی فراهم آورده است، مکانیسم داروینیسم با اطمینان خاطر بنا نهاده میشود و انسان باید این را درک کند که تنها یک تصادف محض است.»[10]
اما وجود کد غشایی نشان میدهد که قاعدهی اصلی اشتباه است و این ایدهی ماتریالیستی که فرگشت هدایت نشده است، نه میتواند پیچیدگی مشخص اطلاعات DNA را توجیه کند و نه اطلاعات مشخص در کد غشایی را که بسیار پیچیدهتر از آن است. همان طور که اطلاعات موجود در DNA دلالت بر طراحی میکنند، اطلاعات آن سوی DNA هم این چنیناند.
- William A. Dembski, “Intelligent design as a theory of information,” February 20, 1997.
- Stephen C. Meyer, “DNA by design,” Rhetoric and Public Affairs 1 (1998): 519-556.
- Roger A. Laine, “A calculation of all possible oligosaccharide isomers both branched and linear yields 1.05 x 1012 structures for a reducing hexasaccharide,” Glycobiology 4 (1994): 759-767.
- Hans-Joachim Gabius, “Biological information transfer beyond the genetic code: The sugar code,” Naturwissenschaften 87 (2000): 108-121.
- Jonathan Wells, “Membrane patterns carry ontogenetic information that is specified independently of DNA,” Bio-Complexity 2014 (2): 1-28.
- Riyi Shi and Richard B. Borgens, “Three-dimensional gradients of voltage during development of the nervous system as invisible coordinates for the establishment of embryonic pattern,” Developmental Dynamics 202 (1995): 101-114.
- AiSun Tseng and Michael Levin, “Cracking the bioelectric code: Probing endogenous ionic controls of pattern formation,” Communicative and Integrative Biology 6 (2013): e22595
- Robert O. Poyton, “Memory and membranes: The expression of genetic and spatial memory during the assembly of organelle macrocompartments,” Modern Cell Biology 2 (1983): 15-72.
- Thomas Cavalier-Smith, “The membranome and membrane heredity in development and evolution,” pp. 335-351 in Robert P. Hirt and David S. Horner (editors), Organelles, Genomes and Eukaryote Phylogeny (Boca Raton, FL: CRC Press, 2004), 348.
- Jacques Monod, quoted in Horace Freeland Judson, The Eighth Day of Creation: The Makers of the Revolution in Biology (New York: Simon & Schuster, 1979), 217.
منبع:
https://evolutionnews.org/2018/06/life-exponential-life-exhibits-intelligent-design-at-many-levels/
[1] William Dembski
[2] Stephen Meyer
[3] Riyi Shi
[4] Richards Borgens
[5] AiSun Tseng
[6] Michael Levin
[7] Robert Poyton
[8] Thomas Cavalier-Smith
[9] James Watson
[10] Francis Crick
[11] Jacques Monod



