
تکامل و مشکل واسطههای غیرکارکردیکیسی لاسکین/ ترجمه: فاطمه بوالحسنی

عدم کارکرد و پیچیدگیکاهشناپذیر[۱]
چارلز داروین[۲] در کتاب منشا گونهها[۳] میگوید: «اگر بتوان ثابت کرد اندام پیچیدهای وجود دارد که احتمال ندارد با تغییرات متعدد، متوالی و جزیی تشکیل شده باشد، نظریهی من کاملا شکست میخورد». ]۱[
در تکامل، انتخاب طبیعی[۴] تنها ساختارهایی را حفظ میکند که برای ارگانیسم مزیتی ایجاد میکنند. اگر ساختاری کاربرد و عملکرد ندارد، پس هیچ مزیتی ندارد و اتلاف منابع ارگانیسم است و در این صورت کنار گذاشته میشود. داروین میگوید ممکن است ساختارهایی وجود داشته باشند که برای آنها مراحل میانی عملکردی[۵] غیرممکن باشد؛ برای مثال واسطهها کارکردی نداشته باشند. این اساسا همان چالش ساختارهای با پیچیدگیکاهشناپذیر است، یعنی جایی که ساختارهای واسطه، کارکردی نداشته باشند. مایکل بیهی[۶]، زیستشناس آمریکایی، توضیح میدهد:
سیستمی که مطابق معیار داروین است سیستمی است که پیچیدگیکاهشناپذیر را نشان میدهد. منظورم از پیچیدگیکاهشناپذیر سیستم واحدی است که از چندین بخش که باهم تعامل دارند تشکیل شده و این بخشها در عملکرد اصلی سیستم شرکت دارند و حذف هر کدام از بخشها به وقفه در عملکرد سیستم منجر میشود. سیستمهای پیچیدهیکاهشناپذیر را نمیتوان با تغییرات اندک و پیدرپیِ در سیستم ابتدایی[۷] تولید کرد؛ زیرا بنابر تعریف، هر صورتابتدایی[۸] برای سیستم پیچیدهیکاهشناپذیر، غیرکاربردی است. از آنجا که انتخاب طبیعی برای انتخاب، نیاز به کارکرد[۹] دارد، اگر چیزی به عنوان سیستم بیولوژیکی پیچیدهیکاهشناپذیر وجود داشته باشد میبایست به عنوان واحدی یکپارچه درآید تا انتخاب طبیعی بتواند روی آن تاثیر بگذارد. تقریبا همه پذیرفتهاند چنین رویدادی ناگهانی با تدریجگرایی[۱۰] داروین غیرقابلتطبیق است. ]۴[
در نقل قول بالا، بیهی خاطرنشان میکند که کیفیتی بنیادی برای هر سیستم پیچیدهیکاهشناپذیر وجود دارد به اینصورت که «برای هر صورتاولیه، سیستم پیچیدهیکاهشناپذیری که بخشی از آن حذف شده باشد، بنابر تعریف، غیرکاربردی است». ]۴[ بیهی این تعریف را با دقت شرح میدهد:
مسیر تکاملی سیستم پیچیدهیکاهشناپذیر مسیری است که شامل یک یا چند مرحلهی انتخابنشده باشد (یعنی یک یا چند جهش ضروری اما انتخاب نشده[۱۱] داشته باشد). میزان پیچیدگیکاهشناپذیر، تعداد مراحل انتخابنشده در این مسیر است. ]۱۱[
علاوه بر بیهی؟
بیهی همچنین مشکل واسطههای عملکردی را با موشخرمایی که تلاش میکند از بزرگراهی با هزار راه عبور کند، مقایسه میکند. ]۹[ او در چنین احساسی تنها نیست. بسیاری از زیستشناسان مشکل واسطههای غیرعملکردی را چالشی بزرگ برای نظریهی داروین میدانند. سورن لاوتراپ[۱۲]، زیستشناس سوئدی، میگوید:
دلایل رد طرح پیشنهادی داروین زیاد بود اما اول از همه باید توجه کرد بسیاری از تغییرات اولیه نمیتوانند از طریق انباشتگی مراحل کوچک به وجود بیایند و حتی اگر بتوانند، انتخاب طبیعی نمیتواند روی آنها اثر بگذارد زیرا مراحل اولیه و میانی، بهصرفه نیستند. ] ۲[
رابرت کارول[۱۳]، دیرینهشناس مشهور تکاملی مهرهداران[۱۴]، میپرسد: آیا فرایندهای تدریجی خردفرگشت[۱۵] میتواند ساختارهای پیچیده را تکامل دهد:
آیا تغییرات در خصوصیات فردی، مانند فراوانی نسبی ژنها برای رنگ روشن و تیرهی بال در پروانههای سازگار با آلودگی صنعتی، بهسادگی در طول زمان تکثیر میشوند تا منشا شبپرهها و پروانهها در حشرات، منشا حشرات از بندپایان بدوی[۱۶] یا منشا بندپایان از میان موجودات چندسلولی اولیه در نظر گرفته شوند؟ چهگونه میتوانیم تکامل تدریجی ساختارهای کاملا جدید، مانند بالهای خفاشها، پرندگان و پروانهها را توضیح دهیم، در حالی که تصور عملکرد یک بال نیمهتکاملیافته تقریبا غیرممکن است؟ ]۱۰[
برخی زیستشناسان برای غلبه بر مشکلات واسطههای غیرکارکردی، ماکروموتاسیونها [۱۷]یا جهشهای ناگهانی[۱۸] را پیشنهاد کردهاند که میتوانند موجودات کاملا متفاوتی تولید کنند. استفان جی گولد[۱۹] طرفدار این نظریه نیست اما خاطرنشان میکند:
فقدان شواهد فسیلی برای مراحل میانی(بین تکاملی) بین انتقالهای اصلی در طراحی ارگانیک، در حقیقت به ناتوانی ما حتی در تخیل برای ساخت واسطههای عملکردی انجامیده که در بسیاری موارد به مشکلی دایمی و آزاردهنده برای روایت تدریجیگرایی تکامل منتهی شده است. ]۸[
کسانی که پیشنهاد میکردند جهشهای بزرگ نادر، هیولاهای امیدوار[۲۰] تولید میکنند که ممکن است بعضی از آنها در واقع مزیت بزرگی داشته باشند، مورد استقبال زیستشناسان قرار نگرفتند. دیرینهشناسانی همچون داگلاس اروین و جیمز ولنتاین[۲۱] دلیلش را اینگونه توضیح میدهند:
جهشهای زنده با اثرات ساختاری یا فیزیولوژیکی بالا، بسیار نادر و معمولا بیحاصل هستند. به نظر میرسد شانس دو فرد جهشیافتهی نادر همسان که در نزدیکی کافی برای تولیدمثل قرار داشته باشند بسیار کم است که بتوان آن را رویدادی تکاملی در نظر گرفت. مشکلاتی از این دست برای هیولاهای امیدوار قابلدوام، این تبیینها را غیرقابلدفاع میکند. ]۳[
اروین و ولنتاین این موضوع را در خصوص منشا پلانهای اصلی بدن[۲۲] (شاخه[۲۳]) و برخی دستههای حیوانات دریایی گفتهاند. با این حال، برخی دیگر دارای ساختارهای غیرقابلتغییر دیگری شدند. انجین کورور[۲۴]، تکاملشناس ترکیهای میگوید:
ویژگی مشترک چشم و بالها این است که تنها در صورتی میتوانند کاربردی باشند که کاملا رشد کرده باشند. به عبارت دیگر، یک چشم نیمهتوسعهیافته نمیتواند ببیند، پرندهای با بالهای نیمهشکلگرفته نمیتواند پرواز کند. چهگونگی به وجود آمدن این اندامها یکی از اسرار طبیعت باقیمانده است که نیاز به کشف دارد. (منبع ۵)
مشارکت[۲۵]
گولد برای غلبه بر این مشکل، «پیشسازگاری»[۲۶] را پیشنهاد میکند، یعنی ساختاری که برای یک عملکرد (کاربردی و اختصاصی) بود، بهطور ناگهانی برای عملکردی دیگر (کاملا متفاوت) استفاده میشود. او میگوید:
اگر پرها ابتدا به عنوان عایق و بعدها برای به دام انداختن حشرات به عنوان طعمه عمل میکردند، ممکن است یک بال اولیه بدون هیچ قصدی برای پرواز ساخته شده باشد. ]۸[
اما چرا باید انتظار داشته باشیم که حشرهگیری تصادفی بهنوعی برای تبدیلشدن به اندامی مفید برای پرواز مناسب باشد؟ «پیشسازگاری» یا «مشارکت» این واقعیت را نادیده میگیرند که بالهایی که عملکرد دارند بسیار پیچیدهتر از آنهایی هستند که برای تلهگذاشتن برای حشرات هستند، این وضعیت برای هر سناریو دیگری از این دست هم صادق است: مثلا احتمال این که ساختار والد مشخصی برای انجام برخی عملکردهای دختر[۲۷] نامشخصِ کاملا متفاوت از والد که پیچیدگی بیولوژیکی بالاتری دارد، سودمند باشد بسیار اندک است. با این حال، زیستشناسان تکاملی روش توضیح گولد را بهطور گسترده حتی در سطح مولکولی، مورد استناد قرار میدهند. آکادمی ملی علوم[۲۸] نوشته است:
ساختارها و فرایندهایی که ادعا میشود بهطور «کاهشناپذیر» پیچیدهاند، معمولا دقیق بررسی نشدهاند . . . تکامل سیستمهای پیچیده میتواند به روشهای گوناگونی رخ دهد. انتخاب طبیعی میتواند یکباره بخشهایی از هر سیستمی را برای یک عملکرد کنار هم قرار دهد و سپس، در زمان بعدی، آن بخشها را با دیگر مولفههای سیستم ترکیب کند تا سیستمی تولید شود که عملکرد متفاوتی دارد. ژنها میتوانند از طریق انتخاب طبیعی تکثیر، تغییر و سپس تقویت شوند. ]۱۴[
مکانیسم «همانندسازی ژن»[۲۹] توضیحی رایج برای منشا پیچیدگی بیولوژیکی است. مزیت ادعاشده این است که ژن مضاعفشده[۳۰]، یک نسخه را برای انجام عملکرد اصلی و دیگری را به عنوان مادهی آزمایشی برای انجام کاری جدید ارایه میدهد. با این حال، همانندسازی ژن بسیار نادر است. میانگین ژنهای مضاعفشده تنها یکی در هر صدمیلیون سال است ]۱۲[ و حتی پس از آن، «اکثریت قریببهاتفاق ژنهای مضاعفشده طی چند میلیون سال خاموش میشوند و شمار معدودی از آنها باقی میمانند که متعاقبا تصفیهی(انتخاب منفی) قویای را تجربه میکنند.» ]۱۲[
مطالعهی دیگری نشان داد ژنها بههیچوجه برای جهشیافتن آزاد نیستند و فشار انتخاب شدیدی روی آنها وجود دارد. ]۱۳[ مکانیسمهای اصلیای که از طریق آنها همانندسازی ژن به تکامل کمک میکند، بهخوبی درک نشدهاند:
با این حال، مشخص نیست چهگونه ژنهای مضاعفشده با موفقیت سیر تکاملی را از حالت اولیهی افزونگی کامل[۳۱] که در آن یک نسخه از ژن احتمالا قابلاستفاده است به وضعیت پایداری که در آن انتخاب طبیعی هر دو نسخه را حفظ میکند، هدایت میکنند. همچنین مشخص نیست چهگونه اغلب این وقایع رخ میدهد. ]۱۲[
نکتهی پایانی این است که تبیین جزییات همانندسازی ژن هنوز بهطور دقیق مشخص نیست و این مسیر قطعا بهطور تجربی تایید نشده است. این تبیین که معمولا تنها بر اساس همولوژی پروتیین[۳۲] است، مکانیسم قابلاعتمادی ندارد و کمی بهتر از نادیدهگرفتن آن است.
جهشهای هاکس[۳۳] یا جهشهای معجزه؟
برخی زیستشناسان جهشهای خاصی را در ژنهای هومئوباکس تنظیمی[۳۴] یا «Hox» متصور شدهاند که در آن جهشهای ساده ممکن است قادر به ایجاد تغییرات بزرگ رشدی در یک ارگانیسم باشند؛ جهشهایی که میتواند باعث ایجاد فنوتایپی کاملا متفاوت شود. با این حال، دستکاری ژنهای Hox (هاکس) کمک چندانی به حل مشکل ایجاد ساختارهای زیستی کاربردی جدید نمیکند؛ زیرا ایجاد تغییرات بزرگ در فنوتایپ بهندرت مفید است. ممکن است جهشهای ژن Hox مکانیسم سادهتری برای ایجاد تغییرات بزرگ باشند اما هنوز درگیر مشکل هیولاهای امیدوار هستند:
مانعی که برای دانشمندان وجود دارد این است که اقتصاد زیرکانهی طبیعت، پیچیدگیهای عظیمی را پنهان میکند. محققان در حال یافتن شواهدی هستند که نشان میدهد ژنهای هاکس و ژنهای هومئوباکس غیرهاکس عوامل مستقل نیستند بلکه اعضای شبکههای ژنتیکی گستردهای هستند که صدها و شاید هزاران ژن دیگر را به هم متصل میکنند. تغییر هر جزء باعث تغییر هزاران مولفهی دیگر نیز میشود؛ هر چند نه لزوما برای بهتر شدن. بنابراین، رویای دستکاری با جعبهابزار طبیعت برای زندهکردن چیزی که دانشمندان آن را «هیولای امیدوار» مینامند -مانند ماهی با پا- احتمالا دستنیافتنی باقی میماند. ]۶[
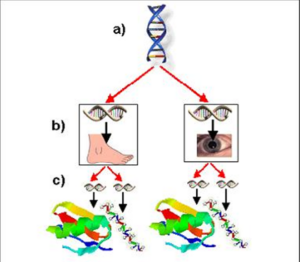
این طرح، سیستم کنترل سلسلهمراتبی و بههمپیوسته را در زیستشناسی نشان میدهد. ژنهای کنترلکنندهی اصلی «Hox»، مانند یک ارتش، دستورات زیستشیمیایی را به ژنهای با رتبهی پایینتر ارسال میکنند. ژنهای Hox (a) ژنهای سطوح پایینتر را کنترل میکنند و با کنترل ژنهای «اعضای بدن» که پروتیینهای ساختاری(c) را ایجاد میکنند، شکلگیری ارگانهای فردی (b) را مدیریت میکنند. جهش در ژنهای سطح بالا احتمالا فاجعهآمیز است مگر اینکه با جهشهای ضروری و بسیار بعید در ژنهای سطح پایین همراه باشد. جهش ژنهای سطح پایین تمایل به تخریب عملکردهای بیولوژیکی کلیدی دارد.
افزون بر این، بسیاری از زیستشناسان هنگام فراخوانی جهشهای ژن Hox فراموش میکنند که ژنهای Hox تنها میتوانند قسمتهایی را که قبلا وجود داشتهاند، دوباره مرتب کنند و اصولا نمیتوانند ساختارهای واقعا جدیدی ایجاد کنند. بحث بسیار ساده این است که ژنها را میتوان در دو دسته در نظر گرفت: ژنهای کنترلکنندهی اصلی (ژن هاکس)[۳۵] و ژنهای اعضای بدن[۳۶].
ژنهای اعضای بدن برای اعضای واقعی بدن کد میکنند در حالی که ژنهای کنترلکنندهی اصلی به ژنهای اعضای بدن میگویند که چه زمانی و کجا بیان شوند و قسمت مربوطهی خود را ایجاد کنند. با این حال، جهشهای هاکس هرگز ژنهای اعضای بدن جدیدی را ایجاد نمیکنند و بنابراین نمیتوانند عملکردهای فنوتایپی واقعا جدیدی را به ژنوم اضافه کنند. در این صورت، در بهترین حالت، ما با مشکلات مرتبط با «پیشسازگاری» مواجه میشویم.
اکثریت تغییرات تکاملی باید از طریق تکامل ژنهای اعضای بدنِ جدید اتفاق بیفتد که جهشهای هاکس نمیتوانند این فرایند را انجام دهند. یکی از منتقدان در مجلهی نیچر[۳۷] این نکته را تشخیص میدهد:
شوارتز[۳۸] این واقعیت را نادیده میگیرد که ژن های هومئوباکس، ژنهای انتخابکننده هستند. اگر ژنهایی که خود به کمک آنها تنظیم میشوند وجود نداشته باشند، نمیتوانند کاری انجام دهند. این ژنها هستند که ساختار تطبیقی اندامها را با جزییات مشخص میکنند. مطمئنا روشنکردن ژن هومئوباکس در مکان نامناسب تنها در صورتی که ژنهای آن اندام در همان فرد وجود داشته باشد میتواند به ظاهرشدن اندامی نابهجا یا اکتوپیک[۳۹] منجر شود. این کاملا اشتباه است که بگوییم یک ماکروموتاسیون میتواند یک چشم را ایجاد کند در حالی که هیچ چشمی در تبار قبلی وجود نداشته است (یعنی این گزاره که ایجاد هر اندامی از طریق ماکروماسیون میتواند اتفاق بیفتد بدون اینکه نیازی به وجود آن اندام در نسلهای گذشته باشد، اشتباه است). جهشهای هومئوتیک[۴۰] اجزا را تغییر میدهند و گاهی ممکن است به تثبیت نوآوریهای تکاملی واقعی منجر شده باشند اما این بدان معنا نیست که چنین تغییراتی در اکثر گونهها وجود دارد. در واقع، ماکروموتاسیونهای این نوع احتمالا اغلب ناسازگار هستند (برخلاف شمار زیادی از گونههای گذشته و حال) و نیازی به ذکر این واقعیت نیست که تفاوتهای مورفولوژیکی میان گونههای مرتبط میتواند بسیار جزیی باشد. ]۷[
جاناتان ولز[۴۱]، زیست شناس، موضوع جهشهای هاکس را در کتاب خود با عنوان نمادهای تکامل[۴۲] مورد بحث قرار میدهد؛ او تشخیص میدهد که ژنهای هاکس را میتوان دستکاری کرد تا پاهای مگس میوهی معمولی (مگس سرکه) از سر آنها جوانه بزنند. سه جهش خاص برای ایجاد این مگس میوهی جهشیافته لازم است و پاها عملکردی ندارند و برای ارگانیسم مفید نیستند. این مثالی عالی از این است که چرا جهشهای معنادار هاکس در ایجاد تغییرات بیولوژیکی بزرگ، پیچیدهتر و کمتر از آنچه که بسیاری وعده دادهاند، ساده هستند و اینکه چهگونه فنوتایپ حاصل معمولا بیفایده و مضر است.
از آنجا که درک این موضوع نسبتا آسان است، ارایهی چند مثال از مورفولوژیهای خرد و کلان که نمیتوانند واسطههای کاربردی داشته باشند مفید است. این نمونهها از نظریهی تکامل تدریجی داروین سرپیچی میکنند و به نظر میرسد سطحی از پیچیدگی را در خود جای دادهاند که دستکم میتوانند اشارهای قوی بر وجود طراح هوشمندی به عنوان علت وجودیشان باشند. سیستمهای بیولوژیکی که واسطههای کاربردی برای آنها غیرممکن به نظر میرسد:
– مکانیزمهای دفاعی در پروانهی ابوالهول
– الزامات شناختی و فیزیولوژیکی برای گفتار انسان
–واسطههای غیرعملکردی در فیزیولوژی انسان
منبع:
http://www.ideacenter.org/contentmgr/showdetails.php/id/841
یادداشتها:
- Origin of the Species by Charles Darwin
2. Lovtrup, S. [professional biologist specialising in Systematics and Developmental Biology, Dept. Animal Physiology, University of Umee, Sweden (also headed the organization of Swedish Developmental Biologists from 1979-87] (1987), Darwinism: The Refutation of a Myth, Croom Helm Ltd., Beckingham, Kent, p. 275
3. Erwin, D..H., and Valentine, J.W. “‘Hopeful monsters,’ transposons, and the Metazoan radiation”, Proc. Natl. Acad. Sci USA 81:5482-5483, Sept 1984
4. Michael Behe, from “Molecular Machines: Experimental Support for the Design Inference” available at “http://www.arn.org/docs/behe/mb_mm92496.htm“.
5. Engin Korur, “Gozlerin ve Kanatlarin Sirri”(The Mystery of the Eyes and the Wings), Bilim ve Teknik, No 203, October 1984, p. 25.
6. Nash J.M., “Where Do Toes Come From?,” Time, Vol. 146, No. 5, July 31, 1995. Also at “http://www.time.com/time/magazine/archive/1995/950731/950731.science.html”
7. Book review of Sudden Origins: Fossils, Genes, and the Emergence of Species by Jeffrey H. Schwartz (Wiley: 1999). by Eors Szathmary in Nature 399:24, June 1999 pg. 745.
8. Stephen Jay Gould (1982), “Is a new and general theroy of evolution emerging?,” In Maynard Smith, J. (ed.), Evolution now A century after Darwin. 129-145. Macmillan Press, London. 239 pp. First published (1980) Paleobiology, 6: 119-130.
9. Darwin’s Black Box by Michael Behe, pg. 141-142.
10. Robert Carroll, Patterns and Processes of Vertebrate Evolution, Cambridge: Cambridge University Press, 1997, pp. 8-10
11. A Response to Critics of Darwin’s Black Box, by Michael Behe, PCID Vol 1.1, Jan/Feb/March 2002; ISCID.org.
12. Lynch, M., Conery, J. S., “The Evolutionary Fate and Consequence of Duplicate Genes” Science 290:۱۱۵۱-۱۱۵۵ (Nov 10, 2000).
13. Huges, Austin L., “Adaptive Evolution of Genes and Genomes”. (see chapter 7, “Evolution of New Protein Function” pp 143-180. (Oxford University Press, New York, 1999).
14. Science and Creationism: A View from the National Academy of Sciences (2nd Ed, 1999; NAP).
[۱]. Irreducible Complexity
[۲]. Charles Darwin
[۳]. Origin of the Species
[۴]. natural selection
[۵]. functional intermediate stages
[۶]. Michael Behe
[۷]. precursor system
[۸] precursor
[۹]. function
[۱۰]. gradualism
[۱۱]. necessary-but-unselected mutations
[۱۲]. Soren Lovtrup
[۱۳]. Robert Carroll
[۱۴]. evolutionist vertebrate paleontologist
[۱۵]. microevolution
[۱۶]. primitive arthropods
[۱۷]. macromutations
[۱۸]. saltations
[۱۹]. Stephen Jay Gould
[۲۰]. hopeful monsters
[۲۱]. Douglas Erwin and James Valentine
[۲۲]. origin of the major body plans of life
[۲۳]. phyla
[۲۴]. Engin Korur
[۲۵]. Co-optation
[۲۶]. pre-adaptation
[۲۷]. daughter function
[۲۸]. The National Academy of Sciences
[۲۹]. gene duplication
[۳۰]. duplicate gene
[۳۱]. initial state of complete redundancy
[۳۲]. protein homology
[۳۳]. Hox-Mutations
[۳۴]. regulatory homeobox genes
[۳۵]. master control genes (Hox genes)
[۳۶]. body part genes
[۳۷] Nature
[۳۸]. Schwartz
[۳۹]. ectopic organ
[۴۰]. Homeotic mutations
[۴۱]. Jonathan Wells



